*Warning scenes and depictions of extreme animal violence will commence
If you are new to this blog or this series of posts you will get best utility by starting with this post and working forwards to follow the thrust of my thought and ideas since these posts build into one another.
All right time to talk about terror birds or, as I like to think of them, the most recent radiation of large bodied terrestrial ziphodont theropods. Yep they have all the hallmarks of a true ziphodont theropod (except for, you know, the teeth); long and muscular neck; strong and rugose orbital brow ridge/top of skull; tall and deep skull braced for strength in the dorso-ventral plane but still relatively lightweight with a central hollow cavity and mechanically weak in the lateral plane; and a relatively weak bite force. Sound like anybody we know?
Now wait a second... what about that whole study pointing to these birds using a unique punch and jab routine in which repeated hammer blows delivered from the tip of the beak killed prey (Degrange et al. 2010)? Well, I am going to be getting into that study on Andalgalornis, in fact the data from that study is what I am going to use to prop up my contention that not only were teratornithids true big game hunting ziphodont predators they were "toothed" but not in the normal sense of the word. Like modern carcass rendering birds (new & old world vultures, petrels) they were equipped with cutlery in the form of a serrated tongue and choanal papillae that interfaced together to commence "choanal grinding" and were true flesh rendering machines of the highest caliber.
Again, note the congruence in general form to other theropods. Deep reinforced skull but with weight saving pneumatic cavities, narrow laterally, vaulted and thick orbital ridge and upper bill to absorb stress and strain. Remind you of somebody else with similar features?
Now Allosaurus had those serrated, mechically reinforced teeth that, I argued, when combined with rapid dorsoventral movements of the neck in a "shimmy" motion instigated a diabolical bone saw technique to saw through skin, muscle, cartilage, and bone. The jaw was weak because the driving power was outsourced to the neck musculature. Phorusrhachids lacked serrated teeth but they did have that strong downcurved beak. It was the hooked tip of the bill that grabbed onto a plug of meat from a prey animal (alive or dead) which brought said plug of meat into the shearing section - the middle of the jaw - I contend. Here a combination of rapid dorsoventral shimmies of the neck and rapid serrated tongue oscillations against the choanal papillae worked to render the meat. After being mechanically compromised, the flesh was broken off with a strong leveraging hook and pull action powered by the entire head, neck, and legs.
![]()
If choanal grinding was the predominant feeding method of these birds we should expect to see the highest capacity for stress and strain along the middle section of the jaw.
![]()
![]()
Granted these various finite element tests (from Degrange et al. 2010) on Andalgalornis, a golden eagle, and cariama were calculated with the bite force (133 N for the terror bird) centered at the tip of the jaw. It is worth looking at where the stress is greatest though. In order to help read this the small chart in the upper left of the diagram goes from relatively weak Von Mises force in the blue upwards to progressively higher forces through the green, blue, yellow, and red colors. If the color is white, well that means the force is off the charts and could potentially imply catastrophic mechanical failure with increasing force. In A, D & G we see that the lateral shake is not looking to good for any of the birds. Note that although the middle of the jaw for Andalgalornis is running red the tip of the beak is off the charts in white realm. I will return to lateral forces later in this post btw. If we look at B, E & H which is a normal bite the whole jaw line is in the nice blue realm for Andalgalornis while the tip of the beak is running red. This suggests that the tip of the beak - which supposedly is meant to withstand the most harrowing pressures when it is slammed into prey - is actually starting to trend into inferior performance during a normal bite pressure. What would happen if it slammed into struggling prey at even higher pressures? And finally we get to the pull back and here again we see the middle of the jaw outperforming the tip of the beak which we can quite readily see is going from red into white color implying the potential for catastrophic failure.
If the tip of the beak really functioned in the battle axe capacity that has been suggested for it as a way to stun/kill prey then we should expect this portion of the skull to show lots of blue color during these strain tests. Except we don't. In all the tests the middle of the jaw - which is where the action is occurring via choanal grinding I suggest - is outperforming the tip of the jaw.
Prediction is met and the battle axe interpretation appears less valid than typical choanal grinding as performed by modern carcass rendering birds (new & old world vultures/petrels) which I first discussed here.
*It should also not go unnoticed, per my previous post theorizing on the mechanical stress relief of theropod head crests/ridges/lacrimal crests that the surparorbital ridges maintain some of the most "blue" areas confirming the mechanical advantage I ascribed them.
![]()
Here are some other problems with the battle axe interpretation:
1)Lack of a modern analogue. If this method of prey acquisition is adaptive we should expect to see at least a few of the 10,000 or so birds in the world evolve some type of analogous prey capture technique. But they are lacking.
2)Difficulty striking a moving, struggling prey item with precision. To kill an animal with a precise blow to the back of the skull or neck as suggested in this method is likely more difficult than assumed. Felids have to grapple and restrain prey in order to initiate their precise biting mechanism. Repeated blows to kill larger prey items seems problematic too as such hooked tips tend to gouge into and get caught in flesh rather than cut through items. We know this due to recent functional tests on the "sickle" claws of dromaeosaurid theropods working more as hooks than slicers. Additionally if a phorusrhacid gets entangled with a large prey item in this manner it is now in danger of sustaining destructive lateral forces from said prey struggling.
3)Dubious depth perception capabilities. For the head to swing back and then into a prey item it is hard to imagine how the phorusrhacid's eyes can stay trained on the prey - especially to hit precise targets such as the back of the skull, vertebral articulation ete etc. If I were to play creator for a second and wanted to design a bird that could pull off this killing technique I would make a sort of composite heron/woodpecker design. Exquisite depth perception, reinforced skull, straight and pointed bill for precise powerful blows. Phorusrhachid skull design is more similar to other carcass rendering birds and extinct theropods and most likely suggests that it fed in a similar fashion.
4)Prey size question. While the battle axe interpretation does not outright dismiss large prey capture it certainly favors a more small/medium game emphasis. This interpretation is ecologically untenable because - with the exception of a few land crocs and marsupial predators - phorusrhacids were far and away the largest, most diverse, and longest lasting predators in South America. They were also the only large South American predators that penetrated into North America. It just makes more ecological sense for them to prey on the largest prey items available to them. No extant terrestrial predators grow to the size of Titanis or Kelenken to primarily hunt prey the size of rabbits or small antelopes.
5)Bite strength. The authors of the Degrange et al. study openly struggle with the relatively weak bite force of Andalgalornis (133 N) which is less than several small mammalian predators. This relatively weak bite force, especially when combined with an increasingly akinetic skull is a bit of an enigma.
However these problems seem to melt away if we invoke a biting adaptation analogous to modern carcass rendering birds (new & old world vultures, giant petrels) and Mesozoic theropods.
![]()
1)Loads of modern analogues for this method. In terror birds what you are essentially looking at is a hopped up lappet-faced vulture or giant petrel. While typically castigated as weak billed scavengers these birds routinely dominate the "predatory" eagles/hawks/falcons at carcasses, engage in combative behavior amongst themselves, drive off and intimidate mammalian carnivores, and are well established hunters and killers in their own right. The absolutely more massive and robust bodied/billed phorusrhachids would be even more impressive in these regards.
2) No precision bite necessary in this model. Prey is bit, hooked into by the bill tip, grinded in the mouth against serrated tongue and choanal papillae, and finally yanked back by the beak/neck & leg musculature until massive trauma facilitates death. Consumption of prey likely began before death. Not pretty or quick but a time honored theropod tradition.
3) Depth perception to a high degree is uncalled for. Through comparison with the way giant petrels and various vultures kill prey - through prolonged and repeated biting, yanking, and grinding encounters - we see it is not necessary for precision bites as called for in the battle axe model i.e. precise depth perception not necessary.
4) Average prey size increases. Giant petrels kill birds almost as large as themselves. Vultures kill relatively large ungulate calves, especially in mobs. Single phorusrhacids were likely capable of killing animals as large, or slightly larger, as themselves and, in groups, targeting animals absolutely larger than themselves. This makes sense ecologically because although South America hosted other large carnivores, phorusrhacids were the most diverse, widespread, long-lasting, and common large predators on the continent. It makes ecological sense for them to be targeting the largest prey available to them. There are no large 50 kg plus terrestrial predators that concentrate or is limited to a prey base that is 1/4 or less of their own size. You might not agree with this assertion but I can't think of one predator that is so ecologically hemmed in. Comments section is open. Peer review has been outsourced to the readers of this post. Generally large carnivores take prey up to and marginally beyond their own size.
5) Strong bite force is not necessary for this model. The cutting of meat via choanal grinding is dependent on speed and friction - not static pressure. The driving musculature facilitating prey trauma and flesh rendering is the serrated tongue and choanal papillae followed by pulls from the neck and legs to pull off bites. These were the two largest muscle masses in the bird's body and are where we should look for possible clues to feeding tactics.
6) Large pneumatic cavity in skull convergent with large foramen (especially antorbital) in ziphodont theropod skulls. Such a weight saving adaptation proves useful in the need for rapid dorsoventral movements of entire head to assist in choanal grinding and pull back motions for prey dismemberment.
There is no need to invoke a feeding method for which there is no modern proxy, and which has significant mechanical and practical problems. A method largely congruent with modern day large carcass rendering birds (new & old world vultures, giant petrels) provides an elegant analogy for phorusrhacid feeding behavior albeit not taken to the extremes that phorusrhacids took such behavior and adaptations. All things being equal, it is more parsimonious to seek modern analogues - especially within the same order - than assume novel and unparalleled feeding adaptations.
For physical confirmation of the likely presence of potentially robust and mechanically reinfornced choanal papillae below is a still taken from the video from this study. You can clearly see the two large ridges of bone - on which the choanal papillae would have sat - in this shot of the roof of the mouth in Andalgalornis.
![]()
![]()
![]()
Now the above gif is not provided to provide simple shock value, shocking as it is, but used to argue a corollary in the feeding apparatus that I argue is largely congruent among phorusrhacids, Mesozoic ziphodont theropods, and modern flesh rendering birds like giant petrels. You should note the strong leveraging capability of both the neck and legs working in conjunction. As well as the rapid "chewing motions" of the beak where I have argued the serrated and stiff tongue is shredding the tissue against the choanal papillae. But more importantly you want to note how the petrel works to keep the fight in the vertical realm. As mentioned earlier and as supported by the Degrange et al. study the phorusrhacid skull is weak when confronted with stresses in the lateral realm. Allosaurus and Mesozoic ziphodont theropods likewise had skulls that were ill-equipped to handle substantial stress in this realm. Petrels and vultures too have skulls likely weak in this realm and their feeding behavior supports this notion. They don't shake their head side to side when interacting with prey or a large carcass. Instead all these animals have a vested interest in keeping the field of battle strictly limited to the vertical plane.
To make a rough analogy to sports let us look at the two main contrasting styles of fighting in mixed martial arts. You have fighters that are most comfortable and effective as upright strikers. They would rather play to their strengths and keep the fight off the mat but upright where their ability for rapid and powerful strikes is most useful. And then you have the specialist floor grapplers. They want to take the fight to the floor where they can utilize their superior grappling skills and specialized submission hold technique. And of course there are fighters that blend the two such as in the pic above where Gina Carano blends grappling with striking to devastating effect in a technique known as "ground and pound". The take home message is that it is in the fighters vested interest to dictate the terms of in what realm the fight occurs based on his/hers respective abilities.
So if we look at the topic from this perspective we can test it to see if it holds any merit based on presumed prey base for these birds. If phorusrhacids were analogous to "strikers" and wanted to keep the fight upright and in the vertical plane we should expect their prey base to be on the whole more like "grapplers" with a lower center of gravity and more geared towards taking the fight to the lateral realm. Which when we look at the presumed prey base of phorusrhacids they are, as a whole, not striking me as especially swift/cursorial track & field superstars.
Notoungulates (including toxodonts): although there are some smaller and swifter varieties like Protypotherium that might have been about as quick as rabbits they are overwhelmingly pretty stolid, clunky bruisers when you get down to it. Certainly no indication that they were on their way to matching the cursorial abilities of derived horses or antelopes. *Update, spoke too soon. Thomas Holtz caught my mistake - there were cursorial herbivores in South America. (see comments below)
Xenarthrans - these are the weird armored guys - including ground/tree sloths, armadillos, glyptodonts, anteaters & tamanduas. Again these animals are not going to win any races.
And later after the great American interchange Gomphotheres were very successful in South America as well.
But don't forget about Macrauchenia. These interesting animals - what is that a camel spliced with an elephant - appear a little leggier than the other critters discussed so far. Speed is always a good tactic but, just giving it an eyeball test I would probably most likely infer it on par with a camel. Probably a good fast trot - but not on par with advanced horses/antelopes.
But, whatever cursorial ability this animal had, what is more important is the suggestion that via its flexible shin and ankle bones these animals had outstanding agility. They could twist and turn on a dime. Admittedly I can't track down an actual source for this ability - it is just repeated several times on the wikipedia page. If anyone has more info, the comments section is open.
All right, so a prey base not necessarily highly cursorial but with armor plated skin, striking hand claws or clubbed tails, large size/robust build, and potentially good maneuverability. Does this sound familar? It should because these characteristics describe the Mesozoic prey base for theropods pretty much tit for tat.
Thyreophoran dinosaurs - stegosaurs, nodosaurs, and ankylosaurs - were both squat, armored, and to varying degrees armed with counterattacking tail clubs, shoulder spikes etc etc. Very reminiscent of xenarthan mammals such as armadillos/glyptodonts.
Sauropods/Saurolophine hadrosaurs - Absolutely larger than theropods but also armed to varying degrees with strong tails/tail clubs, foot claws, dermal armor, and lateral sweeps of neck and tail. Analogous to gomphotheres but also tail wielding glyptodonts and giant sloths to a point.
Therizinosaurs/Giant Oviraptors/Ornithomimids - Hand claws and stout build have often been compared to giant sloths.
Cerartopsids/Pachycephalosaurians aka Marginocephalia - Stout, sturdy build & counterattacking head armament & beaks. Some parallels to notoungulates.
Hadrosaurs/Iguanodonts - Turning agility superior to theropods perhaps analogous to turning ability in Macrauchenia vs. phorusrhacids. Kicks and tails swipes also important.
To have such high congruence in anti-predator strategies in Mesozoic dinosaurs and South American mammals should not be dismissed. I am not the first person to note this congruence. But it is suggestive of a similar method of attack - and weakness - shared between phorusrhacids and Mesozoic predatory theropods.
Faced with theropod marauders you can out-grow, out-fight, out-manoeuvre, or out armor them. Or some combination there of. But none of these methods was likely completely foolproof and phorusrhacids, as their theropod antecedents did likewise, probably were crafty enough to find flaws to exploit in all of the above defenses.
An interesting comparative test is useful when we compare placental mammalian predators/herbivores. The dominant trend here I would argue is that mammalian herbivores have been characterized by an increasing cursorial adaptation. This makes sense when we look at their predators. Canids and hyaenids are pursuit pack hunters. Felids are stealth grapplers. If you are big and strong it is sometimes a good tactic to stand your ground and fight - but this is only viable if you are many orders of magnitude larger and stronger than your predator. Otherwise running is probably the best tactic that works against both felids and canid type predators. Which is taken to extreme levels in various ungulates but especially horses and gazelles. If marsupial predators in South America were the prime danger to the large prey species that lived there we should expect them to evolve primarily in a direction that negated the likely stealth/ambush/grapple strategies employed by borhyaenids / sebecid crocodylomorphs / sabre-toothed Thylacosmilus. Except we don't see that pattern. South American prey species were relatively slow - although sometimes agile - and were more often big, clunky, armored brutes. Exactly opposite the pattern we see in mammalian pack hunting/ambush predator faunas but exactly congruent with Mesozoic ziphodont theropod dominated ecosystems. And like Mesozoic prey species they developed anti-predator armament - tail clubs, large clawed forelimbs, large size - useful in dealing a strong lateral blow to the theropod glass jaw.
The eastern Shasta ground sloth (Nothrotheriops texanus) and the four Titanis walleri terror birds had been at an impasse for several hours. Although both species were recent immigrants to North America the long standing predator prey arms race between these two lineages was not a recent development and heralded back tens of millions of years to their cooevolution on South America. The ground sloth had backed itself into a thicket of palmetto palms and cactus where it turned and faced its attackers.
The terror birds could not attack from the rear here and, faced with the pummeling hand claws of the sloth, were disadvantaged. But the location was in the full hot Florida sun and the birds then commenced a waiting game. When the sloth - after several hours in the sun and facing heat exhaustion - saw several of the scurrilous birds engage in an actual physical fight made it's break for cover. Less than 50 meters away it had excavated an extensive burrow system in the Karst topography. But the sloth would not make it 10 meters.
![]()
Although surprisingly quick, especially in dense brush, the sloth had failed to account for the location of the dominant matriarch of the birds. She, the largest of the flock, stood over 2.5 meters tall and weighed over 170 kg. Having quietly crept down to lie behind some palmettos posterior and adjacent to the sloth she had simply waited for her opportunity. When the sloth made for cover her long legs covered the distance in less than a second. Her aim was deliberate and intentional. She quickly drove into and bit with her hooked beak the softest and most exposed area on the rear of the sloth under the tail - it's anus . And with a series of almost imperceptibly quick serrated tongue and neck oscillations she had established a grip on the sloth's rectum and large intestine. Now with the muscles in her neck and muscular legs working in conjuction she pulled back hard. In a macabre and gruesome tug of war, the sloth pulling in the other direction from the bird helped facilitate its own fatal outcome - disembowelment.
![]()
The sloth realizing it's predicament switched from fleeing to fighting. Mortally wounded it's fate was already sealed. Dancing and lunging from all directions the terror birds harried and harassed the cornered sloth. They knew that exhaustion and blood loss would hasten the death. And with their superior aerobic capacity, speed, and heat threshold the advantage lay firmly with the birds now.
![]()
Perhaps there was good reason for ground sloths to excavate and seek shelter in their extensive tunnel systems....
We can just be thankful for the advent of relatively quick killing predators like felids and constricting/poisonous snakes. The most common method of killing prey during the Mesozoic and in Cenozoic South America was most likely breaching of the body cavity via the anal/cloacal opening. They eat your asshole out of you while you are still alive.
![]()
P.S. Some, or even most, might strongly disagree with my self-publishing style via blog post. Indeed the pitfalls of such method are well documented. I choose this method because it is accessible to all, and - with the use of visuals, video, links, gifs - a superior platform to express ideas in my opinion. Furthermore published academic papers tend to suck the life out of such work, are often behind paywalls, insist on inane formatting rules, cumbersome citation regulations, and peer review is no fool-proof method anyways. To be perfectly transparent I am not going to go through all that. So this is what you get. I know that I have an intelligent and growing audience so I would rather out-source the peer review process to them. If something can survive the slings and arrows of the internet comment section then maybe there is some merit there.
And what does everyone ask as soon as a new paper is published? "Is it online? open access? Can someone send me a pdf?"
Finally going through such prescribed channels is against my D.I.Y. ethos. Why should I PAY some other agency for the privilege to publish MY idea? Especially when they are going to insist on such a dry, cumbersome format? And then THEY get to profit from it?
And to those who simply will not cite, mention, or acknowledge my work because it has not gone through this prescribed process I give you this. If you claim to be a curious, scientifically minded person you should be less concerned with "THROUGH WHAT VENUE I AM GETTING THIS INFORMATION?" but instead "DOES THIS INFORMATION HAVE MERIT?" I have no hidden specimens outside of scientific scrutiny. Whatever I use is from online information available to all.
BTW I have seen popular books, personal communication, unpublished data, anecdotal records all in one form or another cited in reference in published peer reviewed material. So why not blog posts?
Cheers!!
Papers
Degrange FJ, Tambussi CP, Moreno K, Witmer LM, Wroe S (2010) Mechanical Analysis of Feeding Behavior in the Extinct “Terror Bird” Andalgalornis steulleti (Gruiformes: Phorusrhacidae). PLoS ONE 5(8): e11856. doi:10.1371/journal.pone.0011856
![]()
If you are new to this blog or this series of posts you will get best utility by starting with this post and working forwards to follow the thrust of my thought and ideas since these posts build into one another.
All right time to talk about terror birds or, as I like to think of them, the most recent radiation of large bodied terrestrial ziphodont theropods. Yep they have all the hallmarks of a true ziphodont theropod (except for, you know, the teeth); long and muscular neck; strong and rugose orbital brow ridge/top of skull; tall and deep skull braced for strength in the dorso-ventral plane but still relatively lightweight with a central hollow cavity and mechanically weak in the lateral plane; and a relatively weak bite force. Sound like anybody we know?
Now wait a second... what about that whole study pointing to these birds using a unique punch and jab routine in which repeated hammer blows delivered from the tip of the beak killed prey (Degrange et al. 2010)? Well, I am going to be getting into that study on Andalgalornis, in fact the data from that study is what I am going to use to prop up my contention that not only were teratornithids true big game hunting ziphodont predators they were "toothed" but not in the normal sense of the word. Like modern carcass rendering birds (new & old world vultures, petrels) they were equipped with cutlery in the form of a serrated tongue and choanal papillae that interfaced together to commence "choanal grinding" and were true flesh rendering machines of the highest caliber.
| Andalgalornis credit Degrange et al. 2010 |
Again, note the congruence in general form to other theropods. Deep reinforced skull but with weight saving pneumatic cavities, narrow laterally, vaulted and thick orbital ridge and upper bill to absorb stress and strain. Remind you of somebody else with similar features?
 |
| Allosaurus credit Witmer labs |

If choanal grinding was the predominant feeding method of these birds we should expect to see the highest capacity for stress and strain along the middle section of the jaw.

Granted these various finite element tests (from Degrange et al. 2010) on Andalgalornis, a golden eagle, and cariama were calculated with the bite force (133 N for the terror bird) centered at the tip of the jaw. It is worth looking at where the stress is greatest though. In order to help read this the small chart in the upper left of the diagram goes from relatively weak Von Mises force in the blue upwards to progressively higher forces through the green, blue, yellow, and red colors. If the color is white, well that means the force is off the charts and could potentially imply catastrophic mechanical failure with increasing force. In A, D & G we see that the lateral shake is not looking to good for any of the birds. Note that although the middle of the jaw for Andalgalornis is running red the tip of the beak is off the charts in white realm. I will return to lateral forces later in this post btw. If we look at B, E & H which is a normal bite the whole jaw line is in the nice blue realm for Andalgalornis while the tip of the beak is running red. This suggests that the tip of the beak - which supposedly is meant to withstand the most harrowing pressures when it is slammed into prey - is actually starting to trend into inferior performance during a normal bite pressure. What would happen if it slammed into struggling prey at even higher pressures? And finally we get to the pull back and here again we see the middle of the jaw outperforming the tip of the beak which we can quite readily see is going from red into white color implying the potential for catastrophic failure.
 |
| credit Marcos Cenizo |
Prediction is met and the battle axe interpretation appears less valid than typical choanal grinding as performed by modern carcass rendering birds (new & old world vultures/petrels) which I first discussed here.
*It should also not go unnoticed, per my previous post theorizing on the mechanical stress relief of theropod head crests/ridges/lacrimal crests that the surparorbital ridges maintain some of the most "blue" areas confirming the mechanical advantage I ascribed them.

Here are some other problems with the battle axe interpretation:
1)Lack of a modern analogue. If this method of prey acquisition is adaptive we should expect to see at least a few of the 10,000 or so birds in the world evolve some type of analogous prey capture technique. But they are lacking.
2)Difficulty striking a moving, struggling prey item with precision. To kill an animal with a precise blow to the back of the skull or neck as suggested in this method is likely more difficult than assumed. Felids have to grapple and restrain prey in order to initiate their precise biting mechanism. Repeated blows to kill larger prey items seems problematic too as such hooked tips tend to gouge into and get caught in flesh rather than cut through items. We know this due to recent functional tests on the "sickle" claws of dromaeosaurid theropods working more as hooks than slicers. Additionally if a phorusrhacid gets entangled with a large prey item in this manner it is now in danger of sustaining destructive lateral forces from said prey struggling.
3)Dubious depth perception capabilities. For the head to swing back and then into a prey item it is hard to imagine how the phorusrhacid's eyes can stay trained on the prey - especially to hit precise targets such as the back of the skull, vertebral articulation ete etc. If I were to play creator for a second and wanted to design a bird that could pull off this killing technique I would make a sort of composite heron/woodpecker design. Exquisite depth perception, reinforced skull, straight and pointed bill for precise powerful blows. Phorusrhachid skull design is more similar to other carcass rendering birds and extinct theropods and most likely suggests that it fed in a similar fashion.
4)Prey size question. While the battle axe interpretation does not outright dismiss large prey capture it certainly favors a more small/medium game emphasis. This interpretation is ecologically untenable because - with the exception of a few land crocs and marsupial predators - phorusrhacids were far and away the largest, most diverse, and longest lasting predators in South America. They were also the only large South American predators that penetrated into North America. It just makes more ecological sense for them to prey on the largest prey items available to them. No extant terrestrial predators grow to the size of Titanis or Kelenken to primarily hunt prey the size of rabbits or small antelopes.
5)Bite strength. The authors of the Degrange et al. study openly struggle with the relatively weak bite force of Andalgalornis (133 N) which is less than several small mammalian predators. This relatively weak bite force, especially when combined with an increasingly akinetic skull is a bit of an enigma.
Larry Witmer (from here) on bite force:
"We were surprised that the force of biting wasn't greater in Andalgalornis. As we say in the article, our estimate of bite force might be a bit low for some technical reasons, but in general we don't think this bird was delivering hugely powerful bites."
6) Battle Axe/Hammer blow model not consistent with pneumatic structure within skull. Although the skull was large, reinforced mechanically in the dorso-vental plane, and, due to it's size, was absolutely heavy it was made relatively light by a large central pneumatic cavity in the upper jaw. In the battle axe model such a cavity would diminish the impact of blows because of loss of mass. Although, in fairness, such loss of mass could be compensated for by increased acceleration.
 |
| Andalgalornis with special prominence of choanal papillae & serrated tongue (c) Duane Nash |
However these problems seem to melt away if we invoke a biting adaptation analogous to modern carcass rendering birds (new & old world vultures, giant petrels) and Mesozoic theropods.

1)Loads of modern analogues for this method. In terror birds what you are essentially looking at is a hopped up lappet-faced vulture or giant petrel. While typically castigated as weak billed scavengers these birds routinely dominate the "predatory" eagles/hawks/falcons at carcasses, engage in combative behavior amongst themselves, drive off and intimidate mammalian carnivores, and are well established hunters and killers in their own right. The absolutely more massive and robust bodied/billed phorusrhachids would be even more impressive in these regards.
2) No precision bite necessary in this model. Prey is bit, hooked into by the bill tip, grinded in the mouth against serrated tongue and choanal papillae, and finally yanked back by the beak/neck & leg musculature until massive trauma facilitates death. Consumption of prey likely began before death. Not pretty or quick but a time honored theropod tradition.
3) Depth perception to a high degree is uncalled for. Through comparison with the way giant petrels and various vultures kill prey - through prolonged and repeated biting, yanking, and grinding encounters - we see it is not necessary for precision bites as called for in the battle axe model i.e. precise depth perception not necessary.
4) Average prey size increases. Giant petrels kill birds almost as large as themselves. Vultures kill relatively large ungulate calves, especially in mobs. Single phorusrhacids were likely capable of killing animals as large, or slightly larger, as themselves and, in groups, targeting animals absolutely larger than themselves. This makes sense ecologically because although South America hosted other large carnivores, phorusrhacids were the most diverse, widespread, long-lasting, and common large predators on the continent. It makes ecological sense for them to be targeting the largest prey available to them. There are no large 50 kg plus terrestrial predators that concentrate or is limited to a prey base that is 1/4 or less of their own size. You might not agree with this assertion but I can't think of one predator that is so ecologically hemmed in. Comments section is open. Peer review has been outsourced to the readers of this post. Generally large carnivores take prey up to and marginally beyond their own size.
5) Strong bite force is not necessary for this model. The cutting of meat via choanal grinding is dependent on speed and friction - not static pressure. The driving musculature facilitating prey trauma and flesh rendering is the serrated tongue and choanal papillae followed by pulls from the neck and legs to pull off bites. These were the two largest muscle masses in the bird's body and are where we should look for possible clues to feeding tactics.
6) Large pneumatic cavity in skull convergent with large foramen (especially antorbital) in ziphodont theropod skulls. Such a weight saving adaptation proves useful in the need for rapid dorsoventral movements of entire head to assist in choanal grinding and pull back motions for prey dismemberment.
 |
| credit Degrange et al., pneumatic cavity in Andalgalornis skull |
There is no need to invoke a feeding method for which there is no modern proxy, and which has significant mechanical and practical problems. A method largely congruent with modern day large carcass rendering birds (new & old world vultures, giant petrels) provides an elegant analogy for phorusrhacid feeding behavior albeit not taken to the extremes that phorusrhacids took such behavior and adaptations. All things being equal, it is more parsimonious to seek modern analogues - especially within the same order - than assume novel and unparalleled feeding adaptations.
For physical confirmation of the likely presence of potentially robust and mechanically reinfornced choanal papillae below is a still taken from the video from this study. You can clearly see the two large ridges of bone - on which the choanal papillae would have sat - in this shot of the roof of the mouth in Andalgalornis.


 |
| Turkey vulture mouth closeup showing serrated tongue & choanal papillae. credit Williston Conservation Bird Trust Blog |

Now the above gif is not provided to provide simple shock value, shocking as it is, but used to argue a corollary in the feeding apparatus that I argue is largely congruent among phorusrhacids, Mesozoic ziphodont theropods, and modern flesh rendering birds like giant petrels. You should note the strong leveraging capability of both the neck and legs working in conjunction. As well as the rapid "chewing motions" of the beak where I have argued the serrated and stiff tongue is shredding the tissue against the choanal papillae. But more importantly you want to note how the petrel works to keep the fight in the vertical realm. As mentioned earlier and as supported by the Degrange et al. study the phorusrhacid skull is weak when confronted with stresses in the lateral realm. Allosaurus and Mesozoic ziphodont theropods likewise had skulls that were ill-equipped to handle substantial stress in this realm. Petrels and vultures too have skulls likely weak in this realm and their feeding behavior supports this notion. They don't shake their head side to side when interacting with prey or a large carcass. Instead all these animals have a vested interest in keeping the field of battle strictly limited to the vertical plane.
 |
| Gina Carano Ground & Pound CC2.0 |
So if we look at the topic from this perspective we can test it to see if it holds any merit based on presumed prey base for these birds. If phorusrhacids were analogous to "strikers" and wanted to keep the fight upright and in the vertical plane we should expect their prey base to be on the whole more like "grapplers" with a lower center of gravity and more geared towards taking the fight to the lateral realm. Which when we look at the presumed prey base of phorusrhacids they are, as a whole, not striking me as especially swift/cursorial track & field superstars.
Notoungulates (including toxodonts): although there are some smaller and swifter varieties like Protypotherium that might have been about as quick as rabbits they are overwhelmingly pretty stolid, clunky bruisers when you get down to it. Certainly no indication that they were on their way to matching the cursorial abilities of derived horses or antelopes. *Update, spoke too soon. Thomas Holtz caught my mistake - there were cursorial herbivores in South America. (see comments below)
 |
| Homalodotherium cunninghami. Notoungulate. credit Smokeybjb. CC3.0 |
 |
| Nesodon imbricatus. Charles R. Knight. public domain |
Xenarthrans - these are the weird armored guys - including ground/tree sloths, armadillos, glyptodonts, anteaters & tamanduas. Again these animals are not going to win any races.
 |
| Megatherium americanum. Hakan Svensson. CC3.0 |
 |
| Glyptodon & Doedicurus. credit Robert Bruce Horsfall. public domain |
 |
| Stegomastondon mirificus. Wolfman SF. public domain |
 |
| Macrauchenia patachonica. credit Robert Bruce Horsfall. public domain |
All right, so a prey base not necessarily highly cursorial but with armor plated skin, striking hand claws or clubbed tails, large size/robust build, and potentially good maneuverability. Does this sound familar? It should because these characteristics describe the Mesozoic prey base for theropods pretty much tit for tat.
Thyreophoran dinosaurs - stegosaurs, nodosaurs, and ankylosaurs - were both squat, armored, and to varying degrees armed with counterattacking tail clubs, shoulder spikes etc etc. Very reminiscent of xenarthan mammals such as armadillos/glyptodonts.
 |
| Pelorplites & H. sapiens immature female |
 |
| Saurophaganax, Apatosaurus ajax, & Matt Weddel CC4.0 |
Therizinosaurs/Giant Oviraptors/Ornithomimids - Hand claws and stout build have often been compared to giant sloths.
 |
| Nothronychus credit Taylor/Weddel 2013 CC4.0 |
Cerartopsids/Pachycephalosaurians aka Marginocephalia - Stout, sturdy build & counterattacking head armament & beaks. Some parallels to notoungulates.
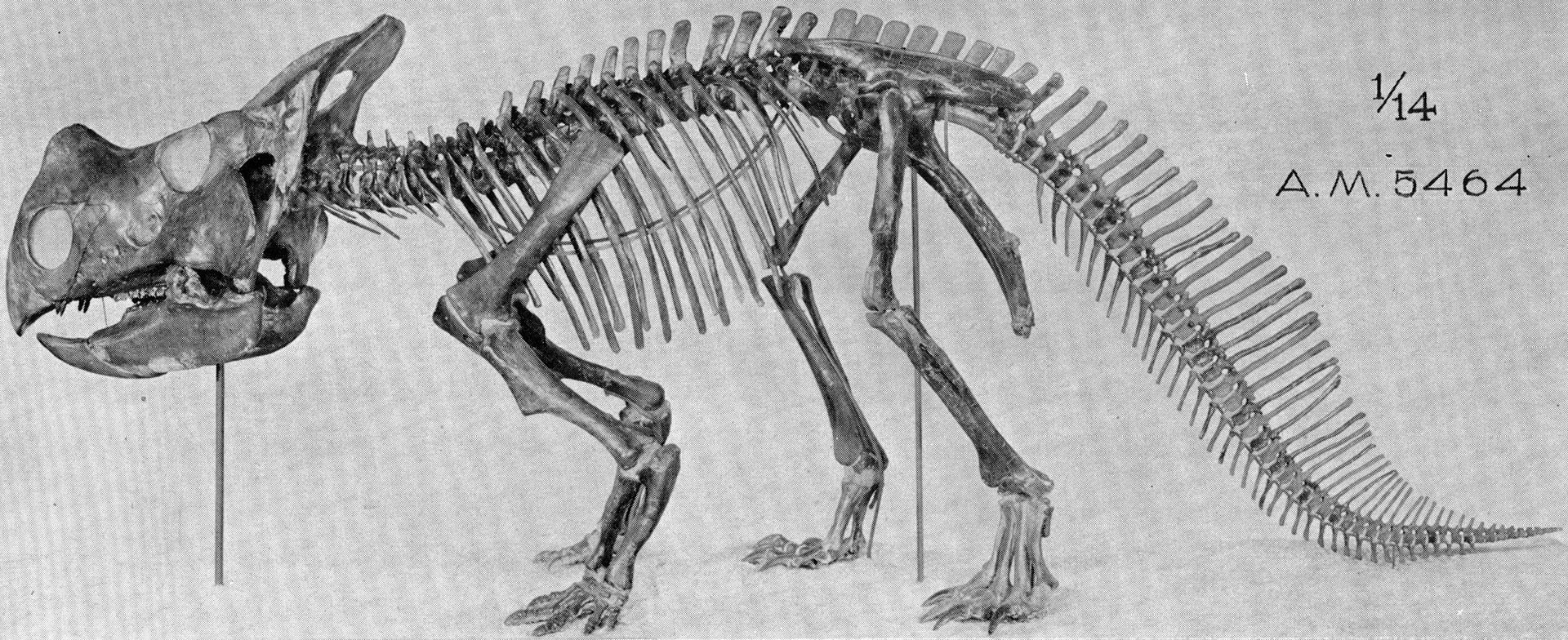 |
| Montanoceratops cerorychos. credit Barnum Brown. public domain |
Hadrosaurs/Iguanodonts - Turning agility superior to theropods perhaps analogous to turning ability in Macrauchenia vs. phorusrhacids. Kicks and tails swipes also important.
 |
| Iguanocolossus. credit lukas Panzarin from PLOS ONE |
To have such high congruence in anti-predator strategies in Mesozoic dinosaurs and South American mammals should not be dismissed. I am not the first person to note this congruence. But it is suggestive of a similar method of attack - and weakness - shared between phorusrhacids and Mesozoic predatory theropods.
Faced with theropod marauders you can out-grow, out-fight, out-manoeuvre, or out armor them. Or some combination there of. But none of these methods was likely completely foolproof and phorusrhacids, as their theropod antecedents did likewise, probably were crafty enough to find flaws to exploit in all of the above defenses.
An interesting comparative test is useful when we compare placental mammalian predators/herbivores. The dominant trend here I would argue is that mammalian herbivores have been characterized by an increasing cursorial adaptation. This makes sense when we look at their predators. Canids and hyaenids are pursuit pack hunters. Felids are stealth grapplers. If you are big and strong it is sometimes a good tactic to stand your ground and fight - but this is only viable if you are many orders of magnitude larger and stronger than your predator. Otherwise running is probably the best tactic that works against both felids and canid type predators. Which is taken to extreme levels in various ungulates but especially horses and gazelles. If marsupial predators in South America were the prime danger to the large prey species that lived there we should expect them to evolve primarily in a direction that negated the likely stealth/ambush/grapple strategies employed by borhyaenids / sebecid crocodylomorphs / sabre-toothed Thylacosmilus. Except we don't see that pattern. South American prey species were relatively slow - although sometimes agile - and were more often big, clunky, armored brutes. Exactly opposite the pattern we see in mammalian pack hunting/ambush predator faunas but exactly congruent with Mesozoic ziphodont theropod dominated ecosystems. And like Mesozoic prey species they developed anti-predator armament - tail clubs, large clawed forelimbs, large size - useful in dealing a strong lateral blow to the theropod glass jaw.
The eastern Shasta ground sloth (Nothrotheriops texanus) and the four Titanis walleri terror birds had been at an impasse for several hours. Although both species were recent immigrants to North America the long standing predator prey arms race between these two lineages was not a recent development and heralded back tens of millions of years to their cooevolution on South America. The ground sloth had backed itself into a thicket of palmetto palms and cactus where it turned and faced its attackers.
 |
| Scrub Palmetto credit Jud McCranie CC4.0 |

Although surprisingly quick, especially in dense brush, the sloth had failed to account for the location of the dominant matriarch of the birds. She, the largest of the flock, stood over 2.5 meters tall and weighed over 170 kg. Having quietly crept down to lie behind some palmettos posterior and adjacent to the sloth she had simply waited for her opportunity. When the sloth made for cover her long legs covered the distance in less than a second. Her aim was deliberate and intentional. She quickly drove into and bit with her hooked beak the softest and most exposed area on the rear of the sloth under the tail - it's anus . And with a series of almost imperceptibly quick serrated tongue and neck oscillations she had established a grip on the sloth's rectum and large intestine. Now with the muscles in her neck and muscular legs working in conjuction she pulled back hard. In a macabre and gruesome tug of war, the sloth pulling in the other direction from the bird helped facilitate its own fatal outcome - disembowelment.

The sloth realizing it's predicament switched from fleeing to fighting. Mortally wounded it's fate was already sealed. Dancing and lunging from all directions the terror birds harried and harassed the cornered sloth. They knew that exhaustion and blood loss would hasten the death. And with their superior aerobic capacity, speed, and heat threshold the advantage lay firmly with the birds now.

Perhaps there was good reason for ground sloths to excavate and seek shelter in their extensive tunnel systems....
 |
| bats in ground sloth paleoburrow Brazil. c/o GeorgiaBeforePeople |
We can just be thankful for the advent of relatively quick killing predators like felids and constricting/poisonous snakes. The most common method of killing prey during the Mesozoic and in Cenozoic South America was most likely breaching of the body cavity via the anal/cloacal opening. They eat your asshole out of you while you are still alive.

P.S. Some, or even most, might strongly disagree with my self-publishing style via blog post. Indeed the pitfalls of such method are well documented. I choose this method because it is accessible to all, and - with the use of visuals, video, links, gifs - a superior platform to express ideas in my opinion. Furthermore published academic papers tend to suck the life out of such work, are often behind paywalls, insist on inane formatting rules, cumbersome citation regulations, and peer review is no fool-proof method anyways. To be perfectly transparent I am not going to go through all that. So this is what you get. I know that I have an intelligent and growing audience so I would rather out-source the peer review process to them. If something can survive the slings and arrows of the internet comment section then maybe there is some merit there.
And what does everyone ask as soon as a new paper is published? "Is it online? open access? Can someone send me a pdf?"
Finally going through such prescribed channels is against my D.I.Y. ethos. Why should I PAY some other agency for the privilege to publish MY idea? Especially when they are going to insist on such a dry, cumbersome format? And then THEY get to profit from it?
And to those who simply will not cite, mention, or acknowledge my work because it has not gone through this prescribed process I give you this. If you claim to be a curious, scientifically minded person you should be less concerned with "THROUGH WHAT VENUE I AM GETTING THIS INFORMATION?" but instead "DOES THIS INFORMATION HAVE MERIT?" I have no hidden specimens outside of scientific scrutiny. Whatever I use is from online information available to all.
BTW I have seen popular books, personal communication, unpublished data, anecdotal records all in one form or another cited in reference in published peer reviewed material. So why not blog posts?
Cheers!!
Papers
Degrange FJ, Tambussi CP, Moreno K, Witmer LM, Wroe S (2010) Mechanical Analysis of Feeding Behavior in the Extinct “Terror Bird” Andalgalornis steulleti (Gruiformes: Phorusrhacidae). PLoS ONE 5(8): e11856. doi:10.1371/journal.pone.0011856
Snively, Cotton, Ridgely & Witmer 2013 Multibody Dynamics Model of Head and Neck Function in Allosaurus (Dinosauria, Theropoda). Paleontologica Electronica May 2013

Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot
Watch me on Deviantart @NashD1. Subscribe to my youtube channel Duane Nash.