Boy I really stepped in it now... Microraptor. For a long time I avoided paraves, Microraptor, the whole dino-bird subject because; there is a lot of talk about maniraptorans on the web already; I enjoy focusing on "meatosaurs" (and still do); and, well, quite literally this whole backwater of quasi dinosaur, quasi bird conundrums literally ruins people. I mean it is a war zone out there folks. Seriously now, academic career types - not just free spirited blogger/idea spinners like yours truly - have gotten their heels so dug in on various sides of the debate it is quite astonishing. The stakes are high in this subject of birdy origins and history will see winners and losers... So why not dive in with Microraptor probably one of the most celebrated, controversial, and perplexing dinosaurs and certainly maniraptorans of the last 25 years? I would definitely put this animal in an unholy triumvirate with Deinonychus and Archaeopteryx in terms of paradigm shifting, scratching your head, game changing iconoclastic status. It really is an odd bird and one we still have loads of unanswered questions about.
So for this piece I am mainly going to side step questions about the aerodynamic efficiency of Microraptor. I don't have a wind tunnel, nor a model to test it in, nor an advanced understanding of aerodynamics. To summarize my current opinion on Microraptor's flight capability I am hedging towards rather limited flight capability - possibly more of a burst flyer of short distances, great maneuverability, with some gliding ability though far from ideal due to all that drag it incurs. I think that the biplane model with the legs spread wide is far from established as I still have questions about ,well, could it anatomically splay its legs? So for now I am going to go with a base line of fairly limited flight capability with pretty good maneuverability and pitch control via the tail and hind wings. I am not wedded to this interpretation and am open to new insight.
I will be working from this point in my attempt at a reasonable lifestyle analysis of this critter. I think it fairly conservative and in line with much of the research into this animal.
Now onto another very contentious issue: terrestrial or arboreal? In order to untangle why this issue is so contentious - I don't think it need be actually - it is worth looking at this question through another lens. The issue of terrestrial versus arboreal in Microraptor has double implications because whether or not this animal was terrestrial or arboreal provides a robust layer of argument to the issue of the competing ground up or trees down hypotheses in the origin of flight in aves proper. And we all know how contentious that issue is and how various factions have their heels dug in on either side of the debate.
The answer to the question of arboreal versus terrestrial habits in Microraptor is quite literally that there is no question of arboreal versus terrestrial in Microraptor's head. And it is in the headspace of Microraptor that we need to get into here - not the head of a tenured professor with a life times' academic stake in the question of ground up or trees down origin of flight. That is quite literally the last head we need to get into in this question. We need to get into the head of a 1 to 2 kilogram predator of small things. An animal that goes recklessly and relentlessly where its prey is. If that prey is roosting 20 meters up a giant old growth ginkgo tree by golly that is where Microraptor is going. If prey is swimming and swarming in the shallows of a lake Microraptor is having a go at that prey. If prey is scurrying from the safety of a burrow in the still of the night Microraptor is right there to greet them. If prey is secreted away deep inside the hollow of fallen tree trunk, Microraptor is sniffing out that quarry and going in that rotten log and making their life hell. Microraptor is kicking ass and taking names and if you are a small swarming, swimming, flying, or scurrying thing and living in the same neighborhood as Microraptor you better watch your back because Microraptoris coming.
"Lock up your children the axeman is coming"
Yes in the trees, in the shallows, in the little cracks and crevices of the environment where the small things hide out Microraptor is going there. If Microraptor could evolve a human intelligence and humour and knew of our debates concerning it Microraptor would be laughing at the pedantic back & forth of terrestrial versus arboreal because Microraptor goes where Microraptor wants to go. Microraptor does not see a sharp division of terrestrial versus arboreal realms. Microraptor sees a 3 dimensional world where the subterranean, aquatic, terrestrial, arboreal, and aerial bleed into one another and Microraptor bleeds into them and exploits them all as it sees fit.
Seeing the world in such a 3 dimensional matrix of positional vortexes is not at all dissonant with how modern supple fingered, clawed, predatory, and small bodied tetrapods exploit and utilize their environment. We see echoes of this multi-functionality in mustelids, varanids, viverids, and probably most intuitively felids, especially our own house cats. I think it was Thomas Holtz who compared microraptors to small felids and I think the comparison is very apt. If you have ever wondered why your house cat has a penchant for seeking high ground - both to hide from threats and survey potential prey - you are seeing echoes of Microraptor in fluffy. I can also recall Darren Naish mentioning in several blog posts that although microraptorines (and other dromies) did not have highly specialized arboreal capabilities that does not mean we should exclude them from scrambling around in the trees, bushes, i.e. exploiting the >shrubbery<.
Of course the bauplan Microraptor started with is different from mammals and lizards. Lacking the low body plan as well as the supple and flexible limbs and torso that allow other tetrapod predators such as lizards and mammals to excel at this particular eco-niche microraptorines evolved alternative solutions to the problems of navigating a cluttered, three dimensional eco-space. The wings on the forearms served to break falls that occurred at elevation, aide in terrestrial balance and pursuit, and provide gliding and flight. Note that I am not concerned with the question of >what< stimulated the evolution of the wings in microraptorines merely how such wings would be useful for a theropod navigating three dimensional space in the pursuit of small game. The >what< of this question (i.e. what caused evolution of wings on the forelimbs) may never be boiled down into a single causal factor but a multiplicity.
Getting back down to earth, literally, I want to address a bit of non-truth that has consistently swirled around microraptorines for a while now. That piece of misinformation - a truism that has always been asserted but never actually demonstrated - is that their "hind-wings" severely hampered terrestrial movement. I see this assertion upheld again and again in both scientific and popular discussions. The idea being presumably that such flight feathers would be a hindrance in locomotion, get caught up in vegetation , and generally cause a kerflunkle for any microraptor trying to just... walk around. That animals with relatively long cursorial looking rear legs; that plot out very much like ground birds morpho-metrically; that this animal could not run or even walk well points to a very mal-adapted and cumbersome beast. Caught in sort of an evolutionary purgatory - not yet an accomplished flyer, glider or climber as well as a cumbersome terrestrial mover - such an animal begs for a quick extinction. Seriously this notion of a terrestrial limited Microraptor should have always have been a non-starter. Kind of like sauropods that could not walk on land. Or giant pterosaurs that could not fly. The kind of "evolutionary experiment" that paleontologists like to posit but can't find any examples of in the real world. Because in the real world their overly cumbersome traits would select them for Darwinian elimination before they even had a statistical chance of entering the fossil record.
Again, we need to get back to what it means to be a predatory small bodied, hot blooded terrestrial tetrapod. You are not just an OKish hunter that sort of blunders through the environment hoping to get lucky enough to snag a meal or two. Such an animal would be selected for extinction quite readily. Nope you are an efficient, ruthless, and relentless hunter. Just look at the pedigree of hunting prowess in small felids and especially small mustelids (i.e. "weasel" types). The true acme of hunting prowess in mammals is not to be found in large canids or felids - they fail all the time. Small bodied predators have to catch a lot more due to their relatively higher metabolism and the relatively small size of their prey. They can't afford to only have a 5% success rate. They are hungry and they are coming. No reason to assume among predatory theropods microraptorines were not the acme of theropod hunting prowess. In fact evolutionary pressure and comparison to modern small bodied tetrapod hunters would dictate that they are.
I mean, come on now, just look at those legs.
Microraptorines could move and move around well on the ground. Assertions that the feathers on the legs would cause undue hardship and toil are simply not meted out when viewed against the panoply of abuse modern birds put their flight feathers through. I was recently watching the flock of California condors that are maintained at the Santa Barbara Zoological Gardens. Splendid birds and you really get a sense for the nuanced social dynamics at play when you watch them for some amount of time. One of the birds sort of crash landed into the branches of a large oak tree in the enclosure after a small skirmish. Mind you this was a California oak (not sure the species) and it had the harsh, spiky leaves of a oak adapted to the drought prone, semi-arid climate of California quite unlike the soft rounded leaves you might encounter on oak leaves in more verdant climes. Anyways, this was no soft landing that the condor made into the tree and it was totally caught up in the brittle, spiky canopy. The bird (second heaviest flying bird in North America) then proceeded to use its wings (e.g. its flight feathers) to literally "swim fly" through the brambly and spiky oak foliage until it could get up to a solid enough limb to perch upon. I got the feeling this was no extraordinary ordeal for the bird and had probably done it a number of times. Not a flight feather lost or even seemingly damaged. Long story short flight feathers - whether on the arm or leg - are not the wimpy structures sometimes assumed to be and are deeply anchored within the dermis or even into the bone and can both receive and deliver substantial abuse and punishment. Microraptorine leg feathers were doubtless anchored well into the leg . Furthermore there is no reason that the feathers on the leg would not have been able to fold up against one another as the leg moved or crouched. It is also entirely possible that such feathers could be moved so as not to scrape against the ground. This is no different than what the flight feathers on the arms of birds and maniraptorans presumably are able to do.
Anyone who argues that bird feathers - especially the pennaceous "flight" feathers whether on the legs or arms - are prone to falling off, getting damaged, or just inhibiting to violent, physical interactions just has not been paying attention to what birds actually do.
Now that we put that whole notion of "could not walk well" to bed for maniraptorines I want to dive a bit deeper into potential uses for the leg feathers. I want to stipulate that these ideas are not mutually exclusive from any of the flight scenarios that people have posited for these animals. Remember that I am not concerned with could or how well Microraptor flew but creating a lifestyle portrait of a small animal killer. Being a specialized small animal killer is an easy inference given the diversity of, well, small animals found in the guts of Microraptor.
To make my case I first want to bracket Microraptor between several other theropods that sported leg feathers. Yes leg feathers were quite en vogue look for several theropods back in the day and not all of them were microraptorines.
Pedopenna daohugouensis. Remember this guy? Yeah I don't either. Just another in the long and confusing line of maniraptorines getting pulled out of the ground in China, I would be lying to you if I don't constantly have to 2x check the names of these guys. Except this species is one that you should take note of. First of all it predates Microraptor and the other Liaoning/Jehol fauna as it dates to somewhere in the mid-late Jurassic (age of the Daohugou beds are debated) but more importantly for our purposes here the legs are feathered - they had hind wings!! The legs are all we really know of this animal and according to wiki: "the long pennaceous feathers of the foot... differ from those of animals like Microraptor. Pedopenna hind wings were smaller and more rounded in shape. The longest feathers were slightly shorter than the metatarsus at about 55 mm (2 in.) long. Additionally, the feathers of Pedopenna were symmetrical, unlike the asymmetrical feathers of some deinonychosaurs (including Microraptor) and modern birds."
The symmetrical feathers of the hind wings in Pedopenna (and Anchiornis btw) are interesting as this suggests that they did not have a use aerodynamically. The hind wings are potentially vestigial and point to a flighted or at least gliding ancestor for Pedopenna (Xu & Zhang, 2005). This invokes the strong potential for "microraptorine" style flyer/gliders going back into the mid-Jurassic - very cool. This also suggests that flight could have evolved, got lost, got regained, and evolved in multiple parallel branches over a fairly long span of time. However I am not quite so sold on Pedopenna and Anchiornis being secondarily flightless. If this was true why would they so quickly lose asymmetrical feathers? Would not asymmetrical feathers still be of use in short glides and other such aerial ventures? I mean, these were not large animals so retention of some aerial ability would still be of use in gliding one would presume?
Time to highlight the seldom mentioned fact that the giant compsognathid Sinocalliopteryx also is noted for non-pennaceous feathers along the back of the metarsus and longer ones on the back of the thigh. These feathers did not form a hind wing nor are compsognathids considered to be on their way to flightedness or secondarily flightless. Yet there was some congruence in evolving feathers along the trailing edge of the lower legs in these disparate animals.
To add a further layer to the enigma of hind wings and lower leg feathers is the amazing microraptor on steroids, Changyuraptor a turkey sized double winged and exceptionally long tailed microraptorine that, despite its size, still points towards being somewhat flight capable or at least gliding.
Although I have my doubts about how long this animal could sustain powered flight, it is always good to remember that wild turkeys and peacocks do fly and they also go into trees as well.
I do think it pays to take a look at these flying large galliforme videos. Notice the seemless transition from an arboreal start in the trees to a short distance flighted phase effortlessly transitioning to an efficient terrestrial phase. Even at this size a turkey sees, interprets, and inhabits its world in a 3-dimensional matrix. I guess this "ground bird" with no obvious arboreal adaptations never got the message that it could not use the trees as it sees fit. Not impressed with flying turkey how about flying peacocks - another "ground bird" that goes where it wants to.
And watch these peacocks in China seemingly sail down en mass from elevation. Reminds me a lot of those turkeys sailing in from the tree tops in the earlier video. Very cool.
We can look at Microraptor not as an isolated hind wing animal but one that slots, or is bracketed in a sense not just phylogenetically but behaviorally, between a number of hind wing or at least feathered leg/metatarsal having theropods. And I think it important to speak towards ecological and behavioral congruity in these theropods because, irrespective of phylogeny, similar lifestyles often confer similar adaptations by convergence.
And the unifying behavioral characteristic that unified microraptorines, Anchiornis, Pedopenna, and Sinocalliopteryx is that they were all terrorists of small game. I have actually discussed Sinocalliopteryx gigas before here. Sinocalliopteryx was obviously not arboreal and it was not flighted, near flighted, or secondarily flightless. But it was really good at snacking on animals that were flighted and/or more inclined towards aerial/arboreal capabilities than it was hence the belly full of dromie leg & enantiornithines. Because of this the authors suggest that it was a stealth hunter, a notion that is hard to dismiss.
Unpacking what it means to be a stealth hunter with a theropod bauplan I believe is key to illuminating what stimulated the evolution of hind wings.
When you are a hunter of small things that means that you have to go to where the small things are at. That implies negotiating tight spaces and deep, dense vegetation. If you are negotiating this eco-morphospace as a small mammal, lizard, or better yet snake - no problem you are already low to the ground with supple legs and torso. But if you have the stiff torso and the tall stiff action limbs of a theropod there is less of an exaptation for maneuvering in such tight places which means there is even more impetus on stealth. A theropod stealthy stalker of small game has to get as close as it can as quietly as it can because whatever you are stalking - chances are that once it detects you it has the better capabilities in navigating tight spaces (or just flying or swimming away). And so imagine the stealthy stalking ability of modern herons but imagine it occurring not while wading but while in deep vegetation. One of the problems such an animal would encounter is curtailing or muffling the disturbance and noise that would occur by walking on and brushing against vegetation/leaf litter/brush with those long, stiff action hind legs. A soft, padded foot would help with muffling footfalls. Likewise a trailing line of feathers along the back of the leg and going down the metatarsus to the foot would potentially muffle or curtail the audible "snapback" of brush that occurs when a leg moves through vegetation - especially dried and brittle twigs, stems etc. etc. Anyone who has done any significant off trail bushwhacking knows what I am talking about. As a tall biped we humans are especially noisy, cumbersome, and just maladapted for stealthy stalking through dense brush. In fact if you are going through dense brush in a group one will often find it necessary to grab a branch or stem that might in fact "snapback" and hit the person following you!! Theropods - which share with us a relatively tall, bipedal build - would face a similar dilemma of "noise pollution" as they moved through dense brush. Having a rearward guard of feathers along the back of the leg and foot would allow vegetation to be "caught" and eased back as the leg stealthily moved through it.
That modern ground birds lack such hind wings or even feathers on the back of their legs may in fact have more to do with a lack of birds engaging in cryptic, stealthy stalking of small game in dense brush. An eco-morph today dominated by mammals, snakes, and lizards.
So here we can draw a rough outline for the impetus and evolution of hind wings - appreciating their import in stealth - that is in fact congruent with what is seen in the fossil record.
A Rough Outline on the Evolution of Hind Wings
I Sinocalliopteryx gigas shows what a theropod with only the barest, incipient stages of this evolution would have. Just simple and small trailing lines of non-pennaceous feathers that assisted in stealthy movement through thick brush while stalking small and agile prey. Coelurosaurs had bigger and more audacious plans in their future so the hind-wing evolution and flight never became more realized in this group.
II Pedopenna, Xiaotingia & Anchiornis. From something similar to Sinocallipoteryx ecologically (albeit dromaeosaurine & smaller) animals like Pedopenna& Anchiornis would arise. The hind wing feathers are now pennaceous although they are not yet flight adapted as they are symmetrical (on both arm & leg wings). However these are true small game hunters of the underbrush as such they capably explored a more three dimensional realization of their environment, certainly more so than the relatively giant Sinocalliopteryx. Although not exquisitely adapted for arboreal life they certainly found themselves up in the trees on occasion. The occasional fall was not an issue as both the arm wings and hind wings would serve as safety nets.
Despite consistent assertions to the contrary Anchironis and other hind-winged microraptorines were excellent terrestrial runners. Arguments against efficient terrestrial running have centered on the lack of leg feathers on modern ground birds. However these arguments have not accounted for a scientific control in terms of behavioral ecology. Big herbivorous ground birds are simply not ecologically comparable to stealthy, small game paravian hunters of dense vegetation. When stealthy hunting in deep vegetation is at a premium a trailing edge of feathers along the back of the leg will often develop in small theropods.
Is there a test for my argument that hind-wings & leg feathers are intimately associated with both stealth and a small game carnivorous lifestyle? Such a paravian would have to be showing a roughly comparable level of flight efficiency to microraptorines with one exception in that it was not a hunter of small game but an omnivore/insectivore/herbivore i.e. stealth was not a critical aspect of its lifestyle.
Turns out the fossil record does provide such a test and it's name Eosinopteryx brevipenna. Eosinopteryx comes from the same time and place as Anchiornis and Brian Switek gave it an excellent summary article here. Personally I think Switek was very prescient in observing how the glut of feathered dino-bird things coming down the pike in last couple of decades has caused a bit of a "numbing" effect. We are not as excited about them as we once were. Switek maintains that specimens like Eosinopteryx might yet have special stories to tell. Eosinopteryx does indeed have a special story to tell I suggest. Eosinopteryx does not have hind wings and it shows a trend indicating decreasing predatory habits. Loss of predatory features includes the lack of a toe claw or even highly recurved foot claws at all and very foreshortened snout with reduced dentition.
![]()
Now granted Eosinopteryx is not especially flighted, in fact the arms have been argued to be especially lacking in the mobility needed for flight. But that does not imply it could not have clambered into trees - as other contemporary paravians that did have hind wings likely did - and that it could not have used the arm-wings as a safety net for falls or gliding. Yet it lacked the hind wings of the contemporary but decidedly more predatory animals like Anchiornis. Anchiornis lacked asymmetrical flight feathers itself and, if flighted, was hardly a good pilot. The main deciding trait that separated Eosinopteryx from hind winged paravians I suggest is diminished predatory habits.
*Update 8/26 I had it pointed out to me via Matt Martyniuk that Sapeornis has some hind wing action going on as well as several other enantiornithines such as those from the new Crato formation: link
Although the hypothesis is not as robust as I constructed it do to these exceptions, it is still possible that a use in stealthy predation was still coopted into the ability to fly with better maneuverability with enhanced hind wings...
III Microraptor. From animals similar to Anchiornis Microraptor likely evolved. Microraptor kept the long hind legs and strong cursorial ability of these animals but improved upon their condition by improving utilization of 3-dimensional space. This revolution was chiefly executed via more efficient flighted ability and true asymmetrical flight feathers on the fore & hind wings. The hind wing feathers still allowed for stealth movement through thick vegetation but were now co opted for aerodynamic purposes.
Hind wings in Microraptor allowed two very important facets of predatory behavior. 1) They allowed for increased maneuverability in the air. 2) They allowed for a more stealth approach in the air.
Point 1 is strongly supported via the studies of Hall & Habib et al. (2012) A strong take home message of the work of Hall & Habib is the concept of a "drag tax" in Microraptor's flight. All of the surface areas that increased maneuverability in Microraptor incurred a consequence in terms of speed. As Microraptor negotiated a very cluttered and complex forest ecosystem such compromise was necessary. And as I will argue further in a future post speed was not at the crux of Microraptor's predatory arsenal - maneuverability and stealth were its chief aims.
Point 2 is a new aspect I want to highlight that will be further embellished in my next post. The hind wing of Microraptor - which I argue is an exaptation co opted by small dromaeosaurines (like Anchiornis) that enabled stealth stalking through thick vegetation - now in the flighted form allowed for an appreciable degree of sound muffling while the leg moved through air.
Swing any long cylindrical object through the air with speed - a bat, a twig, or a leg - and you will get an audible "whoosh" as the air rushes around the cylindrically shaped object and collides together on the rear side. But put a trailing edge of feathers on such an object you will appreciably diminish this said "whooshing" effect. Owls have taken this aspect of stealth approach to the extreme via the sound damping effects of their feathers. I am not suggesting that Microraptor had such capabilities as an owl but the concept remains true to point out. And any such advantage a predator can use, even it only incrementally increases its odds, is still a useful and tactical one.
To restate the angle I am taking - how to figure out how a small theropod bauplan most effiectively operated as a small game hunting animal - let me get back to brass tacks. Microraptor hunted small enantironithines - flighted "birds" that were well ahead of Micraptor in terms of flight adaptations. The superiority of enantiornithine flight speed was no hindrance to the success of Microraptor's predation on them. Because Microraptor was a predator and predators use every advantage that they can garner, Microraptor simply honored the time honored tradition of predators immemorial. It struck in the dark.
Why Microraptor was in fact a nocturnal stealth predator of the utmost capability will be addressed on my next post...
papers
Godefroit, P., Demuynck, H., Dyke, G., Hu, D., Escuillie, F., Claeys, P. 2013.Reduced plumage and flight ability of a new Jurassic paravian theropod from China. Nature Communications. 4, 1394. doi: 10.1038/ncomms2389
Hall, JT, Habib, MB, Hone, DWE, Chiappe, LM. Hindwing function in four winged feathered dinosaurs. (2012) online
Ji, S., Ji, Q., Lu J., and Yuan, C. (2007). "A new giant compsognathid dinosaur with long filamentous integuments from Lower Cretaceous of Northeastern China." Acta Geologica Sinica, 81(1): 8-15
Xu, X. & Zhang, F. (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus". Naturwissenschaften. 92 (4): 173–177. Bibcode:2005NW.....92..173X. doi:10.1007/s00114-004-0604-y. PMID 15685441.
Support me on Patreon.
 |
| closeup cast credit Hiroshi Nishimoto CC 2.0 |
So for this piece I am mainly going to side step questions about the aerodynamic efficiency of Microraptor. I don't have a wind tunnel, nor a model to test it in, nor an advanced understanding of aerodynamics. To summarize my current opinion on Microraptor's flight capability I am hedging towards rather limited flight capability - possibly more of a burst flyer of short distances, great maneuverability, with some gliding ability though far from ideal due to all that drag it incurs. I think that the biplane model with the legs spread wide is far from established as I still have questions about ,well, could it anatomically splay its legs? So for now I am going to go with a base line of fairly limited flight capability with pretty good maneuverability and pitch control via the tail and hind wings. I am not wedded to this interpretation and am open to new insight.
I will be working from this point in my attempt at a reasonable lifestyle analysis of this critter. I think it fairly conservative and in line with much of the research into this animal.
 |
| William Beebes' prescient 1915 "Tetrapteryx" 1915 public domain |
Now onto another very contentious issue: terrestrial or arboreal? In order to untangle why this issue is so contentious - I don't think it need be actually - it is worth looking at this question through another lens. The issue of terrestrial versus arboreal in Microraptor has double implications because whether or not this animal was terrestrial or arboreal provides a robust layer of argument to the issue of the competing ground up or trees down hypotheses in the origin of flight in aves proper. And we all know how contentious that issue is and how various factions have their heels dug in on either side of the debate.
The answer to the question of arboreal versus terrestrial habits in Microraptor is quite literally that there is no question of arboreal versus terrestrial in Microraptor's head. And it is in the headspace of Microraptor that we need to get into here - not the head of a tenured professor with a life times' academic stake in the question of ground up or trees down origin of flight. That is quite literally the last head we need to get into in this question. We need to get into the head of a 1 to 2 kilogram predator of small things. An animal that goes recklessly and relentlessly where its prey is. If that prey is roosting 20 meters up a giant old growth ginkgo tree by golly that is where Microraptor is going. If prey is swimming and swarming in the shallows of a lake Microraptor is having a go at that prey. If prey is scurrying from the safety of a burrow in the still of the night Microraptor is right there to greet them. If prey is secreted away deep inside the hollow of fallen tree trunk, Microraptor is sniffing out that quarry and going in that rotten log and making their life hell. Microraptor is kicking ass and taking names and if you are a small swarming, swimming, flying, or scurrying thing and living in the same neighborhood as Microraptor you better watch your back because Microraptoris coming.
"Lock up your children the axeman is coming"
Yes in the trees, in the shallows, in the little cracks and crevices of the environment where the small things hide out Microraptor is going there. If Microraptor could evolve a human intelligence and humour and knew of our debates concerning it Microraptor would be laughing at the pedantic back & forth of terrestrial versus arboreal because Microraptor goes where Microraptor wants to go. Microraptor does not see a sharp division of terrestrial versus arboreal realms. Microraptor sees a 3 dimensional world where the subterranean, aquatic, terrestrial, arboreal, and aerial bleed into one another and Microraptor bleeds into them and exploits them all as it sees fit.
Seeing the world in such a 3 dimensional matrix of positional vortexes is not at all dissonant with how modern supple fingered, clawed, predatory, and small bodied tetrapods exploit and utilize their environment. We see echoes of this multi-functionality in mustelids, varanids, viverids, and probably most intuitively felids, especially our own house cats. I think it was Thomas Holtz who compared microraptors to small felids and I think the comparison is very apt. If you have ever wondered why your house cat has a penchant for seeking high ground - both to hide from threats and survey potential prey - you are seeing echoes of Microraptor in fluffy. I can also recall Darren Naish mentioning in several blog posts that although microraptorines (and other dromies) did not have highly specialized arboreal capabilities that does not mean we should exclude them from scrambling around in the trees, bushes, i.e. exploiting the >shrubbery<.
Of course the bauplan Microraptor started with is different from mammals and lizards. Lacking the low body plan as well as the supple and flexible limbs and torso that allow other tetrapod predators such as lizards and mammals to excel at this particular eco-niche microraptorines evolved alternative solutions to the problems of navigating a cluttered, three dimensional eco-space. The wings on the forearms served to break falls that occurred at elevation, aide in terrestrial balance and pursuit, and provide gliding and flight. Note that I am not concerned with the question of >what< stimulated the evolution of the wings in microraptorines merely how such wings would be useful for a theropod navigating three dimensional space in the pursuit of small game. The >what< of this question (i.e. what caused evolution of wings on the forelimbs) may never be boiled down into a single causal factor but a multiplicity.
Getting back down to earth, literally, I want to address a bit of non-truth that has consistently swirled around microraptorines for a while now. That piece of misinformation - a truism that has always been asserted but never actually demonstrated - is that their "hind-wings" severely hampered terrestrial movement. I see this assertion upheld again and again in both scientific and popular discussions. The idea being presumably that such flight feathers would be a hindrance in locomotion, get caught up in vegetation , and generally cause a kerflunkle for any microraptor trying to just... walk around. That animals with relatively long cursorial looking rear legs; that plot out very much like ground birds morpho-metrically; that this animal could not run or even walk well points to a very mal-adapted and cumbersome beast. Caught in sort of an evolutionary purgatory - not yet an accomplished flyer, glider or climber as well as a cumbersome terrestrial mover - such an animal begs for a quick extinction. Seriously this notion of a terrestrial limited Microraptor should have always have been a non-starter. Kind of like sauropods that could not walk on land. Or giant pterosaurs that could not fly. The kind of "evolutionary experiment" that paleontologists like to posit but can't find any examples of in the real world. Because in the real world their overly cumbersome traits would select them for Darwinian elimination before they even had a statistical chance of entering the fossil record.
Again, we need to get back to what it means to be a predatory small bodied, hot blooded terrestrial tetrapod. You are not just an OKish hunter that sort of blunders through the environment hoping to get lucky enough to snag a meal or two. Such an animal would be selected for extinction quite readily. Nope you are an efficient, ruthless, and relentless hunter. Just look at the pedigree of hunting prowess in small felids and especially small mustelids (i.e. "weasel" types). The true acme of hunting prowess in mammals is not to be found in large canids or felids - they fail all the time. Small bodied predators have to catch a lot more due to their relatively higher metabolism and the relatively small size of their prey. They can't afford to only have a 5% success rate. They are hungry and they are coming. No reason to assume among predatory theropods microraptorines were not the acme of theropod hunting prowess. In fact evolutionary pressure and comparison to modern small bodied tetrapod hunters would dictate that they are.
I mean, come on now, just look at those legs.
 |
| Specimens by Jaime Headden CC3.0 - http://qilong.deviantart.com/art/Microraptor-Skeletons-27326378 |
Microraptorines could move and move around well on the ground. Assertions that the feathers on the legs would cause undue hardship and toil are simply not meted out when viewed against the panoply of abuse modern birds put their flight feathers through. I was recently watching the flock of California condors that are maintained at the Santa Barbara Zoological Gardens. Splendid birds and you really get a sense for the nuanced social dynamics at play when you watch them for some amount of time. One of the birds sort of crash landed into the branches of a large oak tree in the enclosure after a small skirmish. Mind you this was a California oak (not sure the species) and it had the harsh, spiky leaves of a oak adapted to the drought prone, semi-arid climate of California quite unlike the soft rounded leaves you might encounter on oak leaves in more verdant climes. Anyways, this was no soft landing that the condor made into the tree and it was totally caught up in the brittle, spiky canopy. The bird (second heaviest flying bird in North America) then proceeded to use its wings (e.g. its flight feathers) to literally "swim fly" through the brambly and spiky oak foliage until it could get up to a solid enough limb to perch upon. I got the feeling this was no extraordinary ordeal for the bird and had probably done it a number of times. Not a flight feather lost or even seemingly damaged. Long story short flight feathers - whether on the arm or leg - are not the wimpy structures sometimes assumed to be and are deeply anchored within the dermis or even into the bone and can both receive and deliver substantial abuse and punishment. Microraptorine leg feathers were doubtless anchored well into the leg . Furthermore there is no reason that the feathers on the leg would not have been able to fold up against one another as the leg moved or crouched. It is also entirely possible that such feathers could be moved so as not to scrape against the ground. This is no different than what the flight feathers on the arms of birds and maniraptorans presumably are able to do.
Anyone who argues that bird feathers - especially the pennaceous "flight" feathers whether on the legs or arms - are prone to falling off, getting damaged, or just inhibiting to violent, physical interactions just has not been paying attention to what birds actually do.
Now that we put that whole notion of "could not walk well" to bed for maniraptorines I want to dive a bit deeper into potential uses for the leg feathers. I want to stipulate that these ideas are not mutually exclusive from any of the flight scenarios that people have posited for these animals. Remember that I am not concerned with could or how well Microraptor flew but creating a lifestyle portrait of a small animal killer. Being a specialized small animal killer is an easy inference given the diversity of, well, small animals found in the guts of Microraptor.
To make my case I first want to bracket Microraptor between several other theropods that sported leg feathers. Yes leg feathers were quite en vogue look for several theropods back in the day and not all of them were microraptorines.
Pedopenna daohugouensis. Remember this guy? Yeah I don't either. Just another in the long and confusing line of maniraptorines getting pulled out of the ground in China, I would be lying to you if I don't constantly have to 2x check the names of these guys. Except this species is one that you should take note of. First of all it predates Microraptor and the other Liaoning/Jehol fauna as it dates to somewhere in the mid-late Jurassic (age of the Daohugou beds are debated) but more importantly for our purposes here the legs are feathered - they had hind wings!! The legs are all we really know of this animal and according to wiki: "the long pennaceous feathers of the foot... differ from those of animals like Microraptor. Pedopenna hind wings were smaller and more rounded in shape. The longest feathers were slightly shorter than the metatarsus at about 55 mm (2 in.) long. Additionally, the feathers of Pedopenna were symmetrical, unlike the asymmetrical feathers of some deinonychosaurs (including Microraptor) and modern birds."
The symmetrical feathers of the hind wings in Pedopenna (and Anchiornis btw) are interesting as this suggests that they did not have a use aerodynamically. The hind wings are potentially vestigial and point to a flighted or at least gliding ancestor for Pedopenna (Xu & Zhang, 2005). This invokes the strong potential for "microraptorine" style flyer/gliders going back into the mid-Jurassic - very cool. This also suggests that flight could have evolved, got lost, got regained, and evolved in multiple parallel branches over a fairly long span of time. However I am not quite so sold on Pedopenna and Anchiornis being secondarily flightless. If this was true why would they so quickly lose asymmetrical feathers? Would not asymmetrical feathers still be of use in short glides and other such aerial ventures? I mean, these were not large animals so retention of some aerial ability would still be of use in gliding one would presume?
Time to highlight the seldom mentioned fact that the giant compsognathid Sinocalliopteryx also is noted for non-pennaceous feathers along the back of the metarsus and longer ones on the back of the thigh. These feathers did not form a hind wing nor are compsognathids considered to be on their way to flightedness or secondarily flightless. Yet there was some congruence in evolving feathers along the trailing edge of the lower legs in these disparate animals.
To add a further layer to the enigma of hind wings and lower leg feathers is the amazing microraptor on steroids, Changyuraptor a turkey sized double winged and exceptionally long tailed microraptorine that, despite its size, still points towards being somewhat flight capable or at least gliding.
 |
| turkey sized microraptor Changyuraraptor credit Emily Willoughby CC4.0 |
I do think it pays to take a look at these flying large galliforme videos. Notice the seemless transition from an arboreal start in the trees to a short distance flighted phase effortlessly transitioning to an efficient terrestrial phase. Even at this size a turkey sees, interprets, and inhabits its world in a 3-dimensional matrix. I guess this "ground bird" with no obvious arboreal adaptations never got the message that it could not use the trees as it sees fit. Not impressed with flying turkey how about flying peacocks - another "ground bird" that goes where it wants to.
And watch these peacocks in China seemingly sail down en mass from elevation. Reminds me a lot of those turkeys sailing in from the tree tops in the earlier video. Very cool.
We can look at Microraptor not as an isolated hind wing animal but one that slots, or is bracketed in a sense not just phylogenetically but behaviorally, between a number of hind wing or at least feathered leg/metatarsal having theropods. And I think it important to speak towards ecological and behavioral congruity in these theropods because, irrespective of phylogeny, similar lifestyles often confer similar adaptations by convergence.
And the unifying behavioral characteristic that unified microraptorines, Anchiornis, Pedopenna, and Sinocalliopteryx is that they were all terrorists of small game. I have actually discussed Sinocalliopteryx gigas before here. Sinocalliopteryx was obviously not arboreal and it was not flighted, near flighted, or secondarily flightless. But it was really good at snacking on animals that were flighted and/or more inclined towards aerial/arboreal capabilities than it was hence the belly full of dromie leg & enantiornithines. Because of this the authors suggest that it was a stealth hunter, a notion that is hard to dismiss.
Unpacking what it means to be a stealth hunter with a theropod bauplan I believe is key to illuminating what stimulated the evolution of hind wings.
When you are a hunter of small things that means that you have to go to where the small things are at. That implies negotiating tight spaces and deep, dense vegetation. If you are negotiating this eco-morphospace as a small mammal, lizard, or better yet snake - no problem you are already low to the ground with supple legs and torso. But if you have the stiff torso and the tall stiff action limbs of a theropod there is less of an exaptation for maneuvering in such tight places which means there is even more impetus on stealth. A theropod stealthy stalker of small game has to get as close as it can as quietly as it can because whatever you are stalking - chances are that once it detects you it has the better capabilities in navigating tight spaces (or just flying or swimming away). And so imagine the stealthy stalking ability of modern herons but imagine it occurring not while wading but while in deep vegetation. One of the problems such an animal would encounter is curtailing or muffling the disturbance and noise that would occur by walking on and brushing against vegetation/leaf litter/brush with those long, stiff action hind legs. A soft, padded foot would help with muffling footfalls. Likewise a trailing line of feathers along the back of the leg and going down the metatarsus to the foot would potentially muffle or curtail the audible "snapback" of brush that occurs when a leg moves through vegetation - especially dried and brittle twigs, stems etc. etc. Anyone who has done any significant off trail bushwhacking knows what I am talking about. As a tall biped we humans are especially noisy, cumbersome, and just maladapted for stealthy stalking through dense brush. In fact if you are going through dense brush in a group one will often find it necessary to grab a branch or stem that might in fact "snapback" and hit the person following you!! Theropods - which share with us a relatively tall, bipedal build - would face a similar dilemma of "noise pollution" as they moved through dense brush. Having a rearward guard of feathers along the back of the leg and foot would allow vegetation to be "caught" and eased back as the leg stealthily moved through it.
That modern ground birds lack such hind wings or even feathers on the back of their legs may in fact have more to do with a lack of birds engaging in cryptic, stealthy stalking of small game in dense brush. An eco-morph today dominated by mammals, snakes, and lizards.
So here we can draw a rough outline for the impetus and evolution of hind wings - appreciating their import in stealth - that is in fact congruent with what is seen in the fossil record.
A Rough Outline on the Evolution of Hind Wings
I Sinocalliopteryx gigas shows what a theropod with only the barest, incipient stages of this evolution would have. Just simple and small trailing lines of non-pennaceous feathers that assisted in stealthy movement through thick brush while stalking small and agile prey. Coelurosaurs had bigger and more audacious plans in their future so the hind-wing evolution and flight never became more realized in this group.
II Pedopenna, Xiaotingia & Anchiornis. From something similar to Sinocallipoteryx ecologically (albeit dromaeosaurine & smaller) animals like Pedopenna& Anchiornis would arise. The hind wing feathers are now pennaceous although they are not yet flight adapted as they are symmetrical (on both arm & leg wings). However these are true small game hunters of the underbrush as such they capably explored a more three dimensional realization of their environment, certainly more so than the relatively giant Sinocalliopteryx. Although not exquisitely adapted for arboreal life they certainly found themselves up in the trees on occasion. The occasional fall was not an issue as both the arm wings and hind wings would serve as safety nets.
 |
| Credit Jaime Headden CC3.0 http://qilong.deviantart.com/art/The-Essence-of-Huxley-160904197 |
Despite consistent assertions to the contrary Anchironis and other hind-winged microraptorines were excellent terrestrial runners. Arguments against efficient terrestrial running have centered on the lack of leg feathers on modern ground birds. However these arguments have not accounted for a scientific control in terms of behavioral ecology. Big herbivorous ground birds are simply not ecologically comparable to stealthy, small game paravian hunters of dense vegetation. When stealthy hunting in deep vegetation is at a premium a trailing edge of feathers along the back of the leg will often develop in small theropods.
The merit of a strong hypothesis, or theory, is that predictions can be made and then tested. One such prediction of hind wings being intimately linked to a behavioral ecology of predatory stealth is that such hind wings should not occur in paravians with diminished predatory behavior i.e. omnivory/frugivory/herbivory. So far the prediction I can make holds up in light of the fossil evidence that we have accrued. Hind wings are only found on predaceous paravians and are completely lacking in the groups suspected of more omnivorous/herbivorous inclinations such as caudipterids/enantiornithines/therizinosaurids/oviraptorids etc. etc.
That we have a diverse and well sampled inventory of lifestyles in paravians and it is only the predatory ones that show up with hind wings is an observation that one should keep in their back pockets.
Is there a test for my argument that hind-wings & leg feathers are intimately associated with both stealth and a small game carnivorous lifestyle? Such a paravian would have to be showing a roughly comparable level of flight efficiency to microraptorines with one exception in that it was not a hunter of small game but an omnivore/insectivore/herbivore i.e. stealth was not a critical aspect of its lifestyle.
Turns out the fossil record does provide such a test and it's name Eosinopteryx brevipenna. Eosinopteryx comes from the same time and place as Anchiornis and Brian Switek gave it an excellent summary article here. Personally I think Switek was very prescient in observing how the glut of feathered dino-bird things coming down the pike in last couple of decades has caused a bit of a "numbing" effect. We are not as excited about them as we once were. Switek maintains that specimens like Eosinopteryx might yet have special stories to tell. Eosinopteryx does indeed have a special story to tell I suggest. Eosinopteryx does not have hind wings and it shows a trend indicating decreasing predatory habits. Loss of predatory features includes the lack of a toe claw or even highly recurved foot claws at all and very foreshortened snout with reduced dentition.

Now granted Eosinopteryx is not especially flighted, in fact the arms have been argued to be especially lacking in the mobility needed for flight. But that does not imply it could not have clambered into trees - as other contemporary paravians that did have hind wings likely did - and that it could not have used the arm-wings as a safety net for falls or gliding. Yet it lacked the hind wings of the contemporary but decidedly more predatory animals like Anchiornis. Anchiornis lacked asymmetrical flight feathers itself and, if flighted, was hardly a good pilot. The main deciding trait that separated Eosinopteryx from hind winged paravians I suggest is diminished predatory habits.
*Update 8/26 I had it pointed out to me via Matt Martyniuk that Sapeornis has some hind wing action going on as well as several other enantiornithines such as those from the new Crato formation: link
Although the hypothesis is not as robust as I constructed it do to these exceptions, it is still possible that a use in stealthy predation was still coopted into the ability to fly with better maneuverability with enhanced hind wings...
III Microraptor. From animals similar to Anchiornis Microraptor likely evolved. Microraptor kept the long hind legs and strong cursorial ability of these animals but improved upon their condition by improving utilization of 3-dimensional space. This revolution was chiefly executed via more efficient flighted ability and true asymmetrical flight feathers on the fore & hind wings. The hind wing feathers still allowed for stealth movement through thick vegetation but were now co opted for aerodynamic purposes.
Hind wings in Microraptor allowed two very important facets of predatory behavior. 1) They allowed for increased maneuverability in the air. 2) They allowed for a more stealth approach in the air.
 |
| Microraptor 4 winged flight credit David Krentz |
Point 2 is a new aspect I want to highlight that will be further embellished in my next post. The hind wing of Microraptor - which I argue is an exaptation co opted by small dromaeosaurines (like Anchiornis) that enabled stealth stalking through thick vegetation - now in the flighted form allowed for an appreciable degree of sound muffling while the leg moved through air.
Swing any long cylindrical object through the air with speed - a bat, a twig, or a leg - and you will get an audible "whoosh" as the air rushes around the cylindrically shaped object and collides together on the rear side. But put a trailing edge of feathers on such an object you will appreciably diminish this said "whooshing" effect. Owls have taken this aspect of stealth approach to the extreme via the sound damping effects of their feathers. I am not suggesting that Microraptor had such capabilities as an owl but the concept remains true to point out. And any such advantage a predator can use, even it only incrementally increases its odds, is still a useful and tactical one.
To restate the angle I am taking - how to figure out how a small theropod bauplan most effiectively operated as a small game hunting animal - let me get back to brass tacks. Microraptor hunted small enantironithines - flighted "birds" that were well ahead of Micraptor in terms of flight adaptations. The superiority of enantiornithine flight speed was no hindrance to the success of Microraptor's predation on them. Because Microraptor was a predator and predators use every advantage that they can garner, Microraptor simply honored the time honored tradition of predators immemorial. It struck in the dark.
Why Microraptor was in fact a nocturnal stealth predator of the utmost capability will be addressed on my next post...
 |
| Night Hunting Microraptor credit Robin Liesens |
papers
Godefroit, P., Demuynck, H., Dyke, G., Hu, D., Escuillie, F., Claeys, P. 2013.Reduced plumage and flight ability of a new Jurassic paravian theropod from China. Nature Communications. 4, 1394. doi: 10.1038/ncomms2389
Hall, JT, Habib, MB, Hone, DWE, Chiappe, LM. Hindwing function in four winged feathered dinosaurs. (2012) online
Ji, S., Ji, Q., Lu J., and Yuan, C. (2007). "A new giant compsognathid dinosaur with long filamentous integuments from Lower Cretaceous of Northeastern China." Acta Geologica Sinica, 81(1): 8-15
(2012) Abdominal Contents from Two Large Early Cretaceous Compsognathids (Dinosauria: Theropoda) Demonstrate Feeding on Confuciusornithids and Dromaeosaurids. PLoS ONE 7(8): e44012.doi:10.1371/journal.pone.0044012
Xu, X. & Zhang, F. (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus". Naturwissenschaften. 92 (4): 173–177. Bibcode:2005NW.....92..173X. doi:10.1007/s00114-004-0604-y. PMID 15685441.
"A Long habit of not thinking a thing wrong, gives it a superficial appearance of being right, and raises at first a formidable outcry in defense of custom". Thomas Paine
Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot
Watch me on Deviantart @NashD1. Subscribe to my youtube channel Duane Nash.







.jpg/1920px-Starling_(5503763150).jpg)




















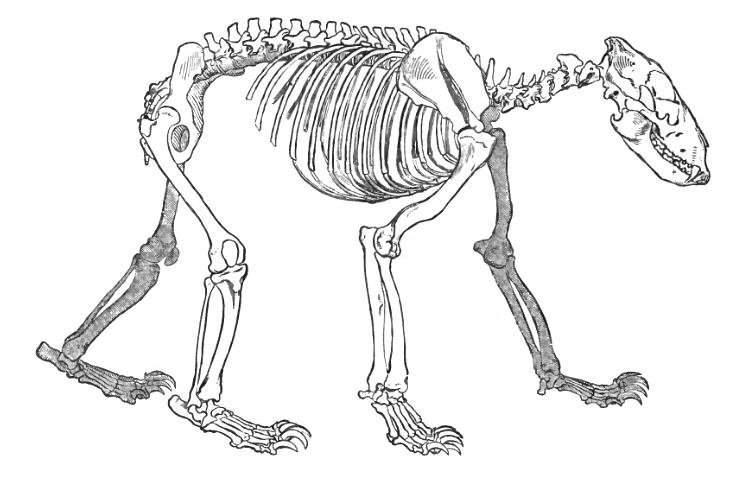
.jpg/800px-Camel_Skeleton_-_Richard_Owen_-_On_the_Anatomy_of_Vertebrates_(1866).jpg)





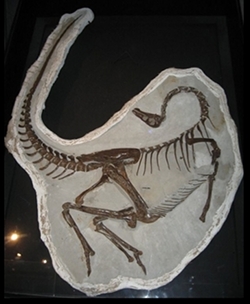




































































_(cropped).jpg)


















.jpg)


_from_Barangay_Dibuluan%2C_San_Mariano_-_ZooKeys-266-001-g069.jpg/800px-Varanus_bitatawa_(KU_322188)_from_Barangay_Dibuluan%2C_San_Mariano_-_ZooKeys-266-001-g069.jpg)




























%2C_Lyme_Regis%2C_England%2C_Early_Jurassic_-_Royal_Ontario_Museum_-_DSC09976.JPG/1280px-Temnodontosaurus_platydon_(cast)%2C_Lyme_Regis%2C_England%2C_Early_Jurassic_-_Royal_Ontario_Museum_-_DSC09976.JPG?1488913210324)
















































































































_(2)_-_geograph.org.uk_-_1342150.jpg/555px-Puffin_(Fratercula_arctica)_(2)_-_geograph.org.uk_-_1342150.jpg)