And now for a change of pace from serrated tongued terror birds and bonesaw theropods to a new look at Masiakasaurus knopfleri in terms of its lifestyle. This is an idea of mine that I have been kicking around for a while so time to dust it off and present it. It's a bit of a stop-gap until I wade back into talking about ziphodont cutlery.
When I look at how an extinct animal likely lived I use a blending of three criteria:
1) Is there a reasonable analogue available? or at least a composite analogue?
2) Is the lifestyle feasible with regards to the local ecology/environment this animal likely lived in? i.e. maybe dedicated piscivory is not the best idea in a semi-arid habitat.
3) Are there any substantiating osteological/biomechanical clues in the skeleton? Trace fossils? Gut contents?
So with Masiakasaurus we have an interesting little character. It's actually the most well known noasaurid. In fact it is one of the better known theropods at this point. Now because Masiakasaurus knopfleri is a relatively recent addition to the theropod canon it gives us a good glimpse of how the suggestion of a lifestyle, then becomes repeated, and then becomes unquestioned. It is in fact a picture perfect view of how dogma arises.
And for Masiakasaurus the lifestyle that is most heavily repeated is the fishing one. Why is this?
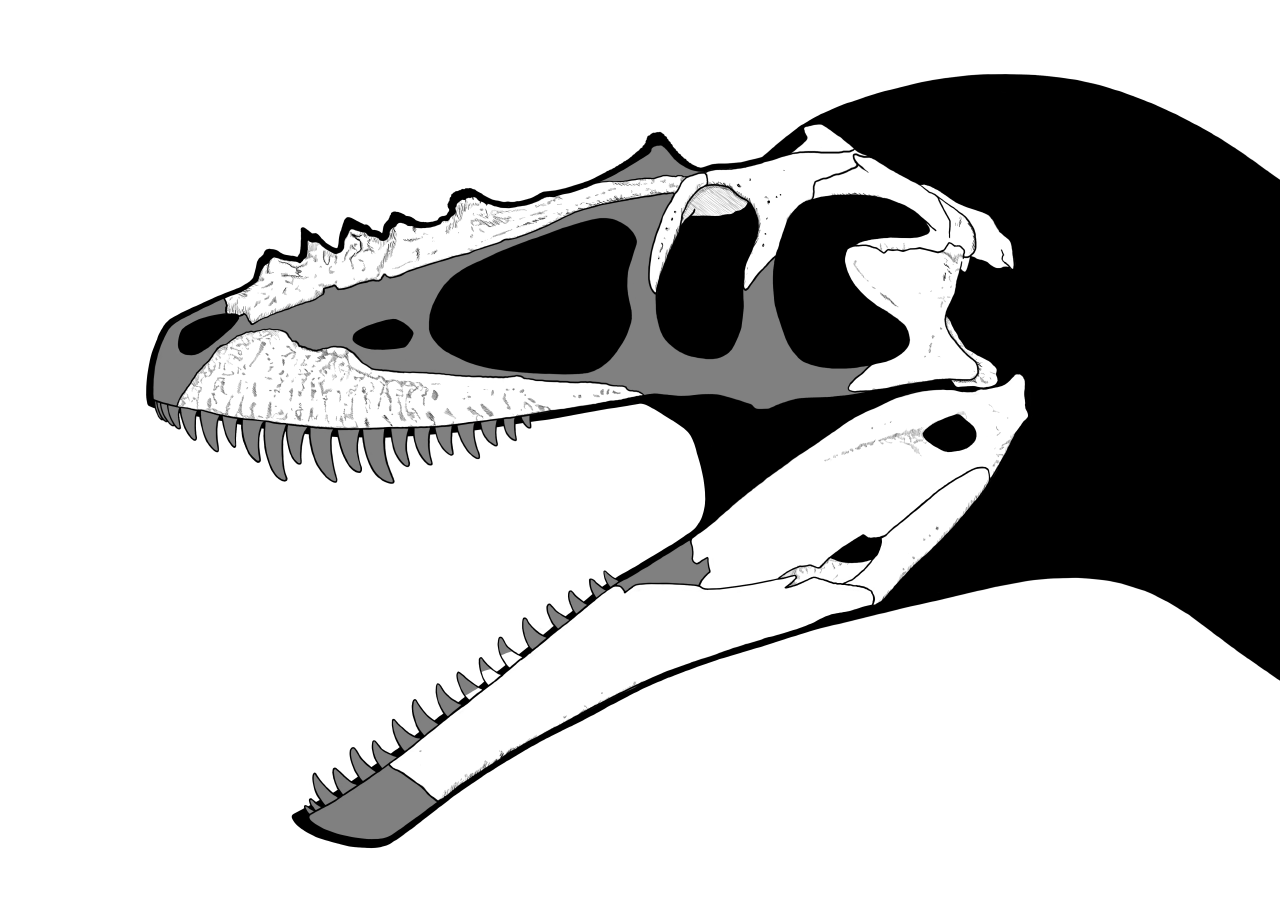
Well first of all let's review why the dental design is so weird in this guy. The anterior end of the jaw is characterized by the procumbent dentition and lack of serrations which grade back into more typical laterally compressed serrated teeth towards the back. Carrano, Sampson & Forster 2002 interpreted this heterodont pattern as indicative of the animal having the ability to grasp at small prey items which could be further minced in the rear teeth. They muse that "One possibility is that Masiakasaurus was insectivorous or piscivorous, using its anterior teeth for acquiring small, whole prey items and its posterior teeth for maceration."
Evolving from this simple suggestion you can clearly trace the insertion of this view as the dominant one with regards to how Masiakasaurus lived. But given that it has really never been fully tested (although Jaime Headden questioned it) is there any merit to dedicated piscivory in Masiakasaurus?
Is fishing Masiakasaurus really just a knee jerk reaction - "Ughhh it's doing something different with those teeth... ummm.... fish!!"
I am going to endorse theirs and others suggestions that the anterior teeth did indeed seize relatively small, single prey items and the posterior teeth did indeed help slice prey items too big to swallow whole. But I don't think that these prey items were commonly fish - which were likely not that common in semiarid Late Cretaceous Madagascar - but that its most common prey was the various likely fossorial animals it shared its island home with.
I hypothesis it was a fossorial animal hunting specialist.
Is there anything in the skeleton of this animal that could substantiate this claim?
If we start working our way back from the head we can see that the neck, at least for a theropod, is relatively straight. And there is an anatomical reason for this. From Carrano 2002: "The posterior cervical vertebrae of Masiakasaurus lack any elevation of the anterior face over the posterior face, suggesting that the base of the neck was relatively straight. They differ from those of coelophysids in being taller and wider relative to their length, bearing larger epipophyses, and having antero-posteriorily shorter neural spines."
To translate the vertebrae did not form the classic sigmoidal "S" shaped neck of theropods. Masiakasaurus was not optimized to achieve the kind of strike that herons do when fishing. Additionally the relatively short vertebrae (taller and wider relative to length) is incongruent with the relatively long vertebrae of azdharchids, herons, and Tanystropheus - all likely strikers of small prey.
So for three tetrapod lineages that - certainly in the case of the heron but most likely in the other two -strike at small prey with their neck in a downward motion and all of which feature very elongated cervical vertebrae, Masiakasaurus does not line up well with.
Additionally from Carrano et al. 2002: "Like abelisaurids and most other basal theropods (but unlike coelurosaurs), the cervical centra zygapophyseal facets are not "flexed" and exposed anteriorly. The three cervicals from the Late Cretaceous Lameta Group of India ascribed to Laevisuchus indicus (Huene & Matley, 1933) are similar to those of Masiakasaurus in most respects, as is the cervical neural arch of Noasaurus. All three display short, anteriorly-placed neural spines, and postzygapophyses that are swept back strongly posteriorily."
What this means, in my shotgun blast translation, is that all that stuff that stuff that sticks out off the cervical centra - the neural spines, the zygapophyses - it is all swept back to streamline the whole contour of the cervical osteology. Almost as if the animal wanted to have any potential obstructions diminished that would impinge it from sticking its neck into tight spots - like deep into a burrow for instance. Furthermore this adaptation might be congruent in other noasaurids hinting at potential similar function and ecology across this group.
An interesting transition occurs when we move from the cervical vertebrae into the dorsal (i.e. torso) vertebrae. Carrano et al. (2011) take note of the interesting transition in this theropod and that it differs from the pattern in most theropods: "The tenth cervical vertebra (C10) lies at or near the cervicodrosal transition, although this can be difficult to define in theropods. Its unusually long proportions indicate that Masiakasaurus was characterized by antero-posteriorly lengthened centra thoughtout the presacral vertebra column. This is unlike the condition in most theropods, where the cervicodorsal transition is marked by one or two anteroposteriorly short vertebrae."
Masiakasaurus is breaking all the rules... but why?
Gone are the short vertebrae of the cervical region instead (from Carrano et al. 2002): "They are spool shaped, weakly amphicoelus, and lack foramina. Unlike the dorsals of Majungatholus, the centra are not shortened but remain anteroposteriorly elongate (approximately twice as long as either as either wide or tall), as in most coelophysids, smaller theropods, and Elaphrosaurus."
When vertebrae are amphicoelus that means both surfaces of a vertebrae are concave and there is good potential for mobility in several directions. This feature, combined with the relatively longer vertebrae, speak to a relatively mobile trunk region. Which is an interesting contrast to the relatively stiffer neck region. Its almost as if the animal needs the ability for its trunk to squeeze and squirm around in tight places - like into a burrow.
![]()
So if the anterior of the body is, as I am suggesting built to get into tight spaces i.e. burrows we should also expect to see some interesting adaptations in the pectoral girdle.
From Carrano et al. 2011: "The scapula and coracoid show an unusual morphology that finds some complacement among other members of Ceratosauria. When articulated, the scapulocoracoid is mediolaterally curved and presents an enormous area for muscle attachment anterior and ventral to the glenoid."
So we have some buffed arms... potentially for digging maybe? However despite the indication of a robust pectoral girdle the morphological theme of trying to "streamline" the bauplan - which I argue is to better access burrows - is persistent.
From Carrano et al. 2011:
"The scapula has a curved blade that reflects the shape of the underlying rib cage, and is broad relative to its length as in coelophysoids as well as other ceratoraurs."
Tranlation: the shoulder blade closely hugs to and mimics the contour of the rib cage. The ecological inference I argue is to diminish obstacles when pummeling into burrows.
"The glenoid has a pronounced anterodorsal rim, as in Ceratosaurus and Majungasarus. In posterior view the scapular portion of the glenoid is D-shaped, and substantially taller dorsoventrally than wide mediolaterally. Its ventral margin along the coracoid suture is oriented obliquely rather than horizontally."
Again the glenoid, or shoulder socket, is showing adaptations to diminish lateral projections and make the anterior of the animals body as flush and streamlined as possible. All useful traits to facilitate entry into tight spaces i.e. burrows.
From Carrano et al. 2011:
"The coracoid is expansive and oval, with long axis oriented anteroposteriorly (fig 18a above). It is much broader than the same element in basal theropods such as Dilophosaurus and Coelophysis, more closely resembling the condition in Elaphrosaurus. Limusaruus, and abelisaurids (although it is more anteroposteriorly elongate than in the latter). The posteroventral process is blunt, projecting only slightly beyond the posteriormost part of the glenoid. It is extremely thin mediolaterally, more so than in almost any other theropod, and lacks the characteristic medial concavity."
Did you pick up on that? The corarcoid, though large, is orientated in a front to back direction but the authors note that it is relatively more elongate than other ceratosaurs. The posteroventral process - a process implies a ridge of bone - is blunt. Not only is it blunted it it is extremely shallow mediolaterally, which means it is thin going from the midline of the body out away from the body - more so than any other theropod the authors mention. Both the bluntness and thiness speak to further diminishing any lateral projection that might diminish fitting the anterior of the body into tight spaces.
Let's look at the humerus now (from Carrono et al 2011):
"Overall it is concave curved medially but nearly straight in the anteroposterior plane. The deltapectoral crest is proportionally short, extending down only about one-third of the total shaft length. The distal condyles, although slightly damaged, are clearly flattened proximodistally as in abelisauruds, Ceratosaurus and Elaphrosaurus. Both the entepicondyle and ectepicondyle are located along the narrow margins close to the distal end, but are indistinctly developed in this specimen."
Now, for our purposes here the most illustrative picture is B, the posterior view - which means you are looking at the humerus from the rear. What you want to notice is that the humerus curves medially- or inwards - from top to bottom. This exquisitely follows the trend that I have been noting all along of lateral projecting bits of anatomy are brought as close in line with the body to precent blocakage into tight spaces. Furthermore the authors note that the condyles - the rugose attachments at the distal end of the humerus - are undeveloped. This feature further streamlines and narrows the anterior aspect of this animal.
But despite these anatomical concessions to diminish lateral projections the pectoral girdle was robust, undiminished and likely very mobile From Carrano et al. 2011:
"No reduction in functionality is evident from the preserved remains. In contrast, the morphology of the humeral head and the expanded muscle origination areas on the ventral pectoral girdle suggest that mobility was significant and perhaps enhanced over the primitive theropod condition."
The morphology of the manual phalanges is interesting - might be useful to compare their morphology with the phalanges of animals that dig....
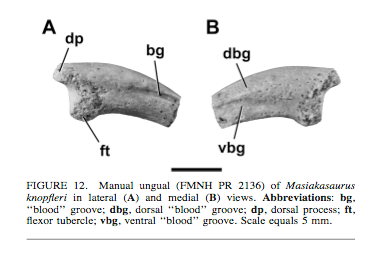
Looking at the manual unguals we see they are relatively short and mediolaterally compressed. They do look a little blunt to be optimized for seizing prey. Could be indicative of some capability in digging.
Moving towards the pelvic area this trend continues.
On the pubic boot (Carranno et al. 2002):
"The distal end is enlarged into a relatively small, rounded "boot" that projects posteriorly form the main shaft axis.... In most other theropods the boot is either small, lobular and unremarkable (coelophysisds), enlarged anteroposteriorly (allosauroids), or lacks an inset (most coelurosaurs)."
Admittedly not the most compelling piece of evidence so far but it is interesting to note the divergence from other theropods and that the "boot" projects posteriorly - again to diminish bony protuberances that might block passage into burrows.
Looking at the femur Carrano et al note:
"The femoral shaft is bowed strongly anteriorly and more subtly medially (fig 14)."
This medial bowing is best seen in B - looking at the femur posteriorly (from the rear). It is quite evident that it bows medially and this feature is consistent with the trend I have been noting of potential laterally projecting bony features being brought close to the body medially. All of which is consistent with a body plan designed to exploit fossorial creatures.
![]()
Does this suggested lifestyle make sense within the paleoecology of where it lived? I must state that here things get a little equivocal as pinpointing true fossorial adaptations in the presumed prey base that I suggest Masiakasaurus exploited is currently lacking.
While evidence of dedicated fossorial specialists that lived with Masiakasaurus is equivocal (but that may change) there are a lot of promising candidates. These are animals that - even if they don't actively burrow themselves - they certainly fall within a size range of animals that frequently use other animals burrows, abandoned or otherwise. Especially in hot arid land with various terrestrial crocs and predatory theropods on the surface.
A diverse fauna of snakes also inhabited Madagascar in the Late Cretaceous. Madstoia madagascariensis is the largest at close to 8 meters and, although no skull is known, was likely a macrophagous predator. Kalyophis, which shows some weakly developed aquatic adaptations. Menarana, which has been suggested to be fossorial. There is a paper online here if you want to read up on them.
Again I am not going to get into the nitty-gritty of were these guys really fossorial. Chances are in a hot arid climate, with predatory crocs and theropods roaming about, and at their size range they likely were. It is certainly a more tenable and viable food base than fish for which I can find no reference to in the Maeverano formation. Furthermore I am not excluding fish (or scavenging, insectivory) as part of their diet. Indeed exploitation of lungfish coiled up in their burrows during the dry season is a very viable option.
And finally I propose Masiakasaurus most analogous to: the Ethiopian Wolf (Canis simensis). This canid of the Ethiopian highlands primarily hunts burrowing rodents - especially big-headed mole rats. And it has specialization in its jaw indicative of this lifestyle with an elongated head, long jaw, widely spaced and slightly procumbent dentition.
So to wrap it up:
The unique and compelling skeletal morphology including; relatively straight neck due to short vertebrae; unique transition to long, flexible dorsal vertebrae from short, stiff neck vertebrae at cervical/dorsal juncture; streamlining of vertebral processes, pectoral and pelvic girdles; potential adaptations for digging in manual unguals/phalanges, mobile pectoral girdle & muscle attachments; medial bowing in of femur & humerus; and long noted procumbent dentition.
A possible diverse and abundant fossorial prey base.
A likely viable and useful modern analogy in the form of the Ethiopian wolf.
All speak to a tenable interpretation of Masiakasaurus knopfleri as a fossorial animal hunting specialist.
Further analysis and discovery of other noasaurid material may show similar adaptations potentially illuminating a Gondwanan radiation of fossorial hunting specialist. Fossorial notosuchians and mammals diversifying at the time may have fostered these specializations.
Can you dig it?
![]()
special thanks to Andrea Cau for providing pdfs of papers
Papers
Carrano, Mathew T., Loewen, Mark A., & Sertich, Joseph J.W. (2011) New materials of Masiakasaurus knopfleri, Sampson, Carrano, and Forster. 2001 and implications for the morphology of the noasauridae (theropoda: ceratosauria). Smithsonia Contributions to Paleobiology number 95
Carrano, Mathew T., Sampson, Scott D., & Forster, Catherine A. (2002) The osteology of Masiakasaurus Knopfleri, a small abelisaurid from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 22(3) Sep 2002 510-534
Sampson, Scott D., Carrano, Mathew T., & Forster, Catherine A. (2001) A bizarre predatory dinosaur from the Late Cretaceous of Madagascar. Nature vol 409. January
![]()
 |
| Masiakaraurus knopfleri credit James St. Johns. CC2.0 |
When I look at how an extinct animal likely lived I use a blending of three criteria:
1) Is there a reasonable analogue available? or at least a composite analogue?
2) Is the lifestyle feasible with regards to the local ecology/environment this animal likely lived in? i.e. maybe dedicated piscivory is not the best idea in a semi-arid habitat.
3) Are there any substantiating osteological/biomechanical clues in the skeleton? Trace fossils? Gut contents?
So with Masiakasaurus we have an interesting little character. It's actually the most well known noasaurid. In fact it is one of the better known theropods at this point. Now because Masiakasaurus knopfleri is a relatively recent addition to the theropod canon it gives us a good glimpse of how the suggestion of a lifestyle, then becomes repeated, and then becomes unquestioned. It is in fact a picture perfect view of how dogma arises.
 |
| used w/permission credit Luis Rey |
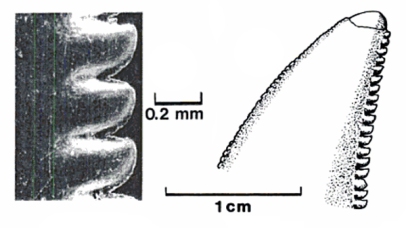
Well first of all let's review why the dental design is so weird in this guy. The anterior end of the jaw is characterized by the procumbent dentition and lack of serrations which grade back into more typical laterally compressed serrated teeth towards the back. Carrano, Sampson & Forster 2002 interpreted this heterodont pattern as indicative of the animal having the ability to grasp at small prey items which could be further minced in the rear teeth. They muse that "One possibility is that Masiakasaurus was insectivorous or piscivorous, using its anterior teeth for acquiring small, whole prey items and its posterior teeth for maceration."
Evolving from this simple suggestion you can clearly trace the insertion of this view as the dominant one with regards to how Masiakasaurus lived. But given that it has really never been fully tested (although Jaime Headden questioned it) is there any merit to dedicated piscivory in Masiakasaurus?
Is fishing Masiakasaurus really just a knee jerk reaction - "Ughhh it's doing something different with those teeth... ummm.... fish!!"
I am going to endorse theirs and others suggestions that the anterior teeth did indeed seize relatively small, single prey items and the posterior teeth did indeed help slice prey items too big to swallow whole. But I don't think that these prey items were commonly fish - which were likely not that common in semiarid Late Cretaceous Madagascar - but that its most common prey was the various likely fossorial animals it shared its island home with.
I hypothesis it was a fossorial animal hunting specialist.
Is there anything in the skeleton of this animal that could substantiate this claim?
 |
| from Carrano et al. 2011 |
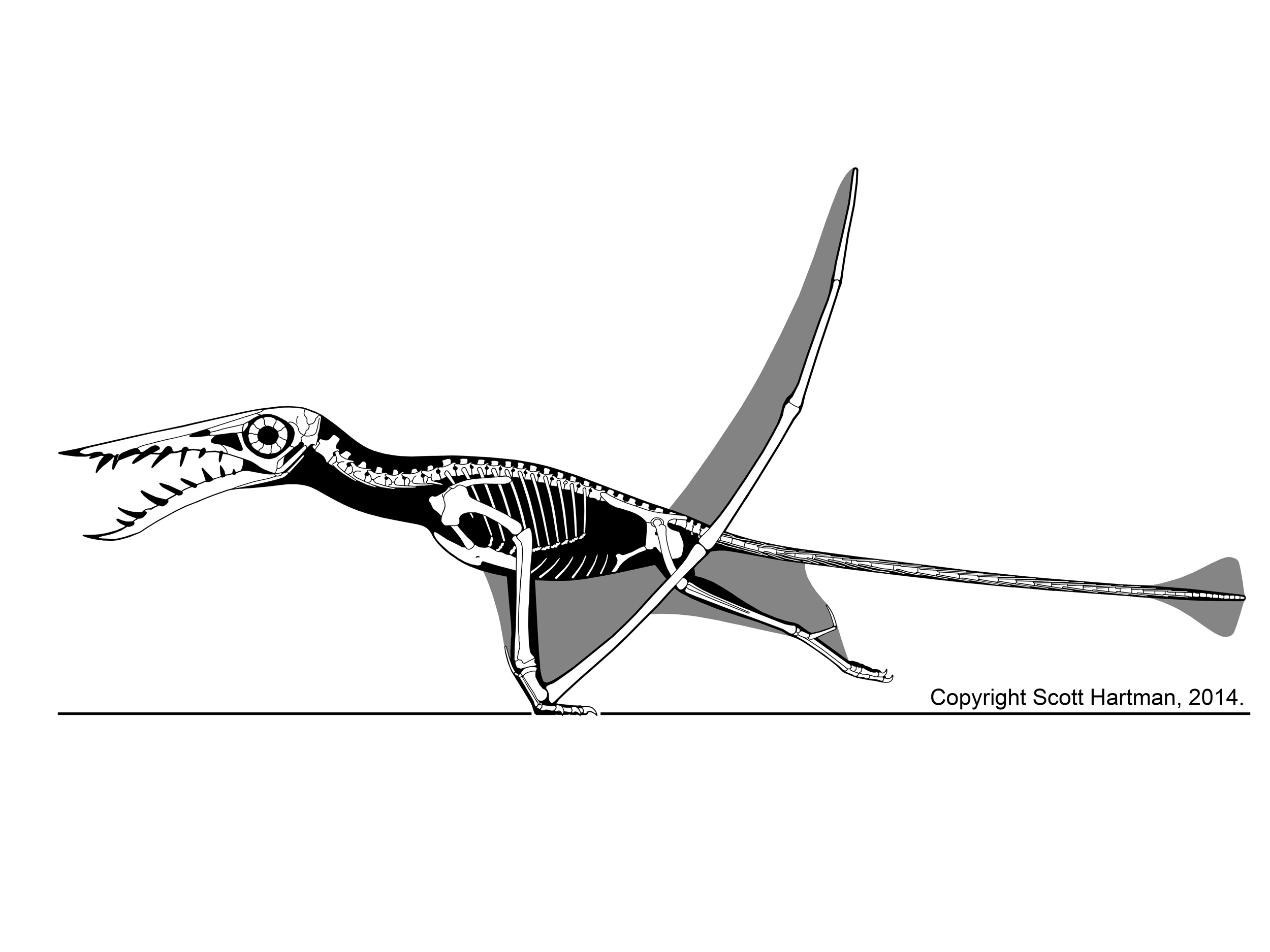
To translate the vertebrae did not form the classic sigmoidal "S" shaped neck of theropods. Masiakasaurus was not optimized to achieve the kind of strike that herons do when fishing. Additionally the relatively short vertebrae (taller and wider relative to length) is incongruent with the relatively long vertebrae of azdharchids, herons, and Tanystropheus - all likely strikers of small prey.
 |
| Quetlzalcoatlus sp. Jaime Headden CC3.0 |
 |
| Tanystropheus longobardicus. CC3.0 credit Ghedoghedo |
 |
| Great Blue Heron. Ardea herodias |
Additionally from Carrano et al. 2002: "Like abelisaurids and most other basal theropods (but unlike coelurosaurs), the cervical centra zygapophyseal facets are not "flexed" and exposed anteriorly. The three cervicals from the Late Cretaceous Lameta Group of India ascribed to Laevisuchus indicus (Huene & Matley, 1933) are similar to those of Masiakasaurus in most respects, as is the cervical neural arch of Noasaurus. All three display short, anteriorly-placed neural spines, and postzygapophyses that are swept back strongly posteriorily."
What this means, in my shotgun blast translation, is that all that stuff that stuff that sticks out off the cervical centra - the neural spines, the zygapophyses - it is all swept back to streamline the whole contour of the cervical osteology. Almost as if the animal wanted to have any potential obstructions diminished that would impinge it from sticking its neck into tight spots - like deep into a burrow for instance. Furthermore this adaptation might be congruent in other noasaurids hinting at potential similar function and ecology across this group.
An interesting transition occurs when we move from the cervical vertebrae into the dorsal (i.e. torso) vertebrae. Carrano et al. (2011) take note of the interesting transition in this theropod and that it differs from the pattern in most theropods: "The tenth cervical vertebra (C10) lies at or near the cervicodrosal transition, although this can be difficult to define in theropods. Its unusually long proportions indicate that Masiakasaurus was characterized by antero-posteriorly lengthened centra thoughtout the presacral vertebra column. This is unlike the condition in most theropods, where the cervicodorsal transition is marked by one or two anteroposteriorly short vertebrae."
Masiakasaurus is breaking all the rules... but why?
Gone are the short vertebrae of the cervical region instead (from Carrano et al. 2002): "They are spool shaped, weakly amphicoelus, and lack foramina. Unlike the dorsals of Majungatholus, the centra are not shortened but remain anteroposteriorly elongate (approximately twice as long as either as either wide or tall), as in most coelophysids, smaller theropods, and Elaphrosaurus."
When vertebrae are amphicoelus that means both surfaces of a vertebrae are concave and there is good potential for mobility in several directions. This feature, combined with the relatively longer vertebrae, speak to a relatively mobile trunk region. Which is an interesting contrast to the relatively stiffer neck region. Its almost as if the animal needs the ability for its trunk to squeeze and squirm around in tight places - like into a burrow.

So if the anterior of the body is, as I am suggesting built to get into tight spaces i.e. burrows we should also expect to see some interesting adaptations in the pectoral girdle.
From Carrano et al. 2011: "The scapula and coracoid show an unusual morphology that finds some complacement among other members of Ceratosauria. When articulated, the scapulocoracoid is mediolaterally curved and presents an enormous area for muscle attachment anterior and ventral to the glenoid."
So we have some buffed arms... potentially for digging maybe? However despite the indication of a robust pectoral girdle the morphological theme of trying to "streamline" the bauplan - which I argue is to better access burrows - is persistent.
From Carrano et al. 2011:
"The scapula has a curved blade that reflects the shape of the underlying rib cage, and is broad relative to its length as in coelophysoids as well as other ceratoraurs."
Tranlation: the shoulder blade closely hugs to and mimics the contour of the rib cage. The ecological inference I argue is to diminish obstacles when pummeling into burrows.
"The glenoid has a pronounced anterodorsal rim, as in Ceratosaurus and Majungasarus. In posterior view the scapular portion of the glenoid is D-shaped, and substantially taller dorsoventrally than wide mediolaterally. Its ventral margin along the coracoid suture is oriented obliquely rather than horizontally."
Again the glenoid, or shoulder socket, is showing adaptations to diminish lateral projections and make the anterior of the animals body as flush and streamlined as possible. All useful traits to facilitate entry into tight spaces i.e. burrows.
 |
| from Carrano et al. 2011 |
From Carrano et al. 2011:
"The coracoid is expansive and oval, with long axis oriented anteroposteriorly (fig 18a above). It is much broader than the same element in basal theropods such as Dilophosaurus and Coelophysis, more closely resembling the condition in Elaphrosaurus. Limusaruus, and abelisaurids (although it is more anteroposteriorly elongate than in the latter). The posteroventral process is blunt, projecting only slightly beyond the posteriormost part of the glenoid. It is extremely thin mediolaterally, more so than in almost any other theropod, and lacks the characteristic medial concavity."
Did you pick up on that? The corarcoid, though large, is orientated in a front to back direction but the authors note that it is relatively more elongate than other ceratosaurs. The posteroventral process - a process implies a ridge of bone - is blunt. Not only is it blunted it it is extremely shallow mediolaterally, which means it is thin going from the midline of the body out away from the body - more so than any other theropod the authors mention. Both the bluntness and thiness speak to further diminishing any lateral projection that might diminish fitting the anterior of the body into tight spaces.
Let's look at the humerus now (from Carrono et al 2011):
"Overall it is concave curved medially but nearly straight in the anteroposterior plane. The deltapectoral crest is proportionally short, extending down only about one-third of the total shaft length. The distal condyles, although slightly damaged, are clearly flattened proximodistally as in abelisauruds, Ceratosaurus and Elaphrosaurus. Both the entepicondyle and ectepicondyle are located along the narrow margins close to the distal end, but are indistinctly developed in this specimen."
 |
| credit Carrano et al. 2011 |
But despite these anatomical concessions to diminish lateral projections the pectoral girdle was robust, undiminished and likely very mobile From Carrano et al. 2011:
"No reduction in functionality is evident from the preserved remains. In contrast, the morphology of the humeral head and the expanded muscle origination areas on the ventral pectoral girdle suggest that mobility was significant and perhaps enhanced over the primitive theropod condition."
The morphology of the manual phalanges is interesting - might be useful to compare their morphology with the phalanges of animals that dig....
 |
| credit Carrano et al. 2011 |
 |
| credit Carrano et al. 2002 |
On the pubic boot (Carranno et al. 2002):
"The distal end is enlarged into a relatively small, rounded "boot" that projects posteriorly form the main shaft axis.... In most other theropods the boot is either small, lobular and unremarkable (coelophysisds), enlarged anteroposteriorly (allosauroids), or lacks an inset (most coelurosaurs)."
Admittedly not the most compelling piece of evidence so far but it is interesting to note the divergence from other theropods and that the "boot" projects posteriorly - again to diminish bony protuberances that might block passage into burrows.
 |
| credit Carrano et al. 2002 |
"The femoral shaft is bowed strongly anteriorly and more subtly medially (fig 14)."
 |
| credit Carrano et al. 2002 |
This medial bowing is best seen in B - looking at the femur posteriorly (from the rear). It is quite evident that it bows medially and this feature is consistent with the trend I have been noting of potential laterally projecting bony features being brought close to the body medially. All of which is consistent with a body plan designed to exploit fossorial creatures.

Does this suggested lifestyle make sense within the paleoecology of where it lived? I must state that here things get a little equivocal as pinpointing true fossorial adaptations in the presumed prey base that I suggest Masiakasaurus exploited is currently lacking.
While evidence of dedicated fossorial specialists that lived with Masiakasaurus is equivocal (but that may change) there are a lot of promising candidates. These are animals that - even if they don't actively burrow themselves - they certainly fall within a size range of animals that frequently use other animals burrows, abandoned or otherwise. Especially in hot arid land with various terrestrial crocs and predatory theropods on the surface.
 |
| Simosuchus clarki, the pug nosed notosuchian. credit Gordon E. Robertson CC3.0 |
 |
| Vintana sertichi credit Lucille Betti-Nash (c) |
 |
| Beelzebufo ampinga. credit Nobu Tamura(spinops.blogspot.com) CC3.0 |
 |
| Araripesuchus patagonicus. remains of the same genus have been recovered from Madagascar. credit Gabriel Lio. CC3.0 |
A diverse fauna of snakes also inhabited Madagascar in the Late Cretaceous. Madstoia madagascariensis is the largest at close to 8 meters and, although no skull is known, was likely a macrophagous predator. Kalyophis, which shows some weakly developed aquatic adaptations. Menarana, which has been suggested to be fossorial. There is a paper online here if you want to read up on them.
Again I am not going to get into the nitty-gritty of were these guys really fossorial. Chances are in a hot arid climate, with predatory crocs and theropods roaming about, and at their size range they likely were. It is certainly a more tenable and viable food base than fish for which I can find no reference to in the Maeverano formation. Furthermore I am not excluding fish (or scavenging, insectivory) as part of their diet. Indeed exploitation of lungfish coiled up in their burrows during the dry season is a very viable option.
 |
| Canis simensis. credit Harri J. CC2.0 |
 |
| Ethiopian Wolf. credti Paul Gervais |
 |
| credit Jaime A. Headden The Strange Case of Dr. Masiaka & Mr. Vicious CC3.0 |
 |
| Duane Nash |
.jpg/1280px-Rare_Ethiopian_Wolf_Feeding%2C_Bale%2C_Ethiopia_(9681610337).jpg) |
| Ethiopian Wolf credit Rod Waddington (lolz) CC2.0 |
So to wrap it up:
The unique and compelling skeletal morphology including; relatively straight neck due to short vertebrae; unique transition to long, flexible dorsal vertebrae from short, stiff neck vertebrae at cervical/dorsal juncture; streamlining of vertebral processes, pectoral and pelvic girdles; potential adaptations for digging in manual unguals/phalanges, mobile pectoral girdle & muscle attachments; medial bowing in of femur & humerus; and long noted procumbent dentition.
A possible diverse and abundant fossorial prey base.
A likely viable and useful modern analogy in the form of the Ethiopian wolf.
All speak to a tenable interpretation of Masiakasaurus knopfleri as a fossorial animal hunting specialist.
Further analysis and discovery of other noasaurid material may show similar adaptations potentially illuminating a Gondwanan radiation of fossorial hunting specialist. Fossorial notosuchians and mammals diversifying at the time may have fostered these specializations.
Can you dig it?

special thanks to Andrea Cau for providing pdfs of papers
Papers
Carrano, Mathew T., Loewen, Mark A., & Sertich, Joseph J.W. (2011) New materials of Masiakasaurus knopfleri, Sampson, Carrano, and Forster. 2001 and implications for the morphology of the noasauridae (theropoda: ceratosauria). Smithsonia Contributions to Paleobiology number 95
Carrano, Mathew T., Sampson, Scott D., & Forster, Catherine A. (2002) The osteology of Masiakasaurus Knopfleri, a small abelisaurid from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 22(3) Sep 2002 510-534
Sampson, Scott D., Carrano, Mathew T., & Forster, Catherine A. (2001) A bizarre predatory dinosaur from the Late Cretaceous of Madagascar. Nature vol 409. January

"A Long habit of not thinking a thing wrong, gives it a superficial appearance of being right, and raises at first a formidable outcry in defense of custom". Thomas Paine
Support me on Patreon.
Support me on Patreon.
Like antediluvian salad on facebook. Visit my other blog southlandbeaver.blogspot
Watch me on Deviantart @NashD1. Subscribe to my youtube channel Duane Nash.



























































































.jpg/1280px-20110721_Berkenhof_Zoo_(0050).jpg)


































































.JPG.png/800px-Machaeroides_eothen_(cropped).JPG.png)










.jpg/1280px-Lions_painting%2C_Chauvet_Cave_(museum_replica).jpg)


































.jpg)







































.jpg/800px-Peregrine_Falcon_(3938274380).jpg)












.JPG/450px-Ursus_thibetanus_3_(Wroclaw_zoo).JPG)


.jpg/800px-Skeksis_on_Display_(red_blur).jpg)